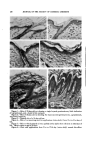
PROGRESS IN THE CHEMISTRY OF DISUI.FIDES 303 this case a helical configuration is impossible and that the only possible configuration under these circumstances is a linear one in which the amino acid groups are as far apart as possible. These facts would satisfactorily account for the normal solubility and normal optical rotation observed for penicillamine disulfide. The marked loss in optical rotatory power involv- ing a change from 220 to 22% a decrease of 90 per cent, resulting from modifying cystine to penicillamine disulfide, enables one to assess the rela- tive contribution that the helical structure itself contributes to the total optical rotation of cystine. Another example which may be cited to illustrate the predominant im- portance of the helical configuration as contributing to the high optical rotatory ability of cystine, may be seen in a comparison with the amino acid cystathionine. Here the structure of cystine is modified such that a methylene group CH2 replaces one of the sulfur atoms. The resulting struc- ture lacks the disulfide linkage and is composed of two identical amino acid groups separated by four atoms consisting of three carbons and one sulfur. Here, too, the helical structure is lacking, and here too, the optical rotation has fallen from 220 ø to a value of 22 ø. An examination of a skeletal molecular model of cystine reveals that three ionic groups (2 carboxylate and one protonated amino groups) form the prongs of the pincer. These lie within a plane forming the apices of a small triangle. External to this triangle and at a considerable distance away is the fourth group (an amino group) of the zwitter ion pair. A pos- tulated structure of this nature satisfactorily explains the anomalous acidic and basic group dissociation constants of cystine, namely: pK• 1.00, pK• 1.7, pK3 7.48, pK4 9.02. It will be noted here immediately that two of these constants are values which are considered to be "normal" for amino acids and two of them are distinctly abnormal. Let us consider the ab- normal dissociation constant pK• which is reported by several workers to be less than unity. This fact is indicative of a very strong acid whose dis- sociation is so great as to be incapable of any precise measurement-- approximately the strength of inorganic acids. The positively charged atmosphere in the vicinity of the carboxylate site is so great as to render it almost impossible for a proton to approach this site and hence the pK is very low. The pK3 of one of the amino groups is also extremely low, that is, it is somewhat reluctant to acquire a proton. This would imply that the proto- hated amine is in a vicinity of negative charges which would be most con- ducive to the release of an acquired proton. V. D•su•.mt)E C•.E^v^oE ^st) T•E C•t•STR¾ OF H^•R W^wso Let us now consider some of the factors operative in disulfide cleavage. It is obvious that one of the factors is the tendency to release of strain im-

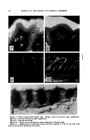
304 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS posed upon a disulfide bond. Prior to our ultraviolet spectral interpreta- tion of disulfide strain there were two methods employed to measure this strain at best these methods were little more than qualitative. The "alkali lability" method used by Clarke (50), Schbberl (51), and others, is based on the time required to produce a precipitate of lead sulfide when a sulfur containing compound is treated with an alkali solution of a soluble lead salt. A second method employed by Aflqeck and Dougherty (52) in the case of cyclic disulfide was to observe the apparent ease with which polymer- ization of the disulfide compound occurred in the presence of AICl:•. In our method not only have we been able to estimate the disulfide strain from ultraviolet spectra analysis, but we have been able to quantitatively inter- pret the spectra in terms of the specific chromophores responsible for the disulfide absorption in the ultraviolet. We have shown that the categori- cal assertion that alkali of itself cleaves the S--S bond is not necessarily true. Many tertiary disulfides are stable in alkali. Actually ultraviolet spectra do not reveal the strain existing in a disulfide bond as such, but rather they reveal the presence of a chromophore. This chromophore is produced as a consequence of the desire on the part of the strained struc- ture to assume a non-strained configuration. The structure invariably assumed by strained disulfides is the conjugated linear form --C•S-- S=C--. This particular chromophoric form will occur in either acid or alkali and depends solely on the structure of the disulfide compound in question. In addition to the apparent strain imposed on the disulfide linkage by pH or structural distortion of the sulfur bond angles, there is another factor affecting S--S cleavage, namely, the steric factor. Here we will attempt to distinguish between internal and external steric factors. What we imply by an internal steric factor may be understood by the following considera- tion. When large or numerous bulky groups are present on both sides of an S--S linkage, these groups will serve to barricade the disulfide against attack by various disulfide-specific reagents. In the case of penicillamine disulfide, reagents which will normally cleave cystine such as sulfite, cya- nide, etc., are now ineffective. This we interpret in terms of the presence of the four methyl groups. Schbberl (53) has similarly indicated in the cases of tetra methyl and tetra phenyl dithiodiglycolic acids, that these com- pounds are not attacked by reagents that ordinarily cleave S--S. Arnold (54) in addition, has reported that tertiary butyl disulfide is extremely diffi- cult to reduce. In all instances cited here bulky groups within the mole- cule are preventing an attack upon the disulfide group. The external steric factor in connection with disulfide cleavage is only manifest in three-dimensional structures such as proteins. The proteins, keratin, cortocin, and insulin, have the common feature that all contain
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)