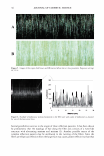
116 JOURNAL OF COSMETIC SCIENCE EVIDENCE AND UTILITY OF MELANIN DEGRADING ENZYMES Thomas Mammone, Ph.D., Ken Marenus, Ph.D., Neelam Muizzuddin and Daniel Maes, Ph.D. Estee Lauder Research Laboratories, 125 Pine/awn Road, Melville, NY 11747 Abstract: The biochemii:al synthesis ofhuDWI melanin is understood in some detail. However, little is known about melanin degradation and catabolism of melanin. We hypothesize that human skin contains enzymes that degrade melanin and these enzymes can be used to reduce skin color. To test this hypothesis, HaCaT keratinocytes and normal human keratinocytes in culture were pulse labeled for one hour with radio labeled s � thetic melanin. 1bis melanin was synthesized in vitro using tyrosinase enzyme from mushrooms and using radiolabeled [ C] 3,4- dihydroxyphenylalanine (DOPA) as a substrate. After the initial pulse labeling. samples of both the cells and media were taken at 2, 4, 6 and 18 hours. Over these time periods the counts remaining in the media and cell fraction were significantly decreased. This data suggests the need for new protein synthesis and the lysosome organelle function for the degradation. Melanin degrading extracts isolated from Aspergillus fumigatus and Saccharomyces cerevisiae were applied to human skin. These extracts cause significant reduction in UVB induced pigmentation. These extracts may be useful in developing new whitening products to even skin color and tone. INTRODUCTION: Previously, Ohtali and Seiji [I) investigated the in vitro degradation of melanosomes and melanin in mouse liver. These authors discovered that melanosomes are degraded by the lysosomes, but that melanin is not appreciably degraded. These experiments were conducted with B 16 mouse melanoma derived melanosomes, which may be more difficult to digest Also mouse liver lysosomes were used and this may not be representative of all tissues. In addition, the duration of incubation with lysosomes was 60 minutes in these experiments and may not have been entirely relevant in the case of melanin in the skin. We have investigated huDWI skin keratinocytes, which would be a physiologically relevant site for a melanin-degrading enzyme. Later investigators reported that lysosomal extracts from huDWI skin were unable to degrade natural melanin [21 and therefore it must be shed intact from the epidermis in squames. These authors also did not observe any changes in melanin injected into guinea pig skin. Although melansomal proteins and lipids did appear to decrease with time in the guinea pig skin. However, we have observed that synthetic radiolabeled melanin, when placed in cultures of human keratinocytes, was lost with increasing time after the initial labeling period. 14CXXXJ 1axm 1axm i IDDl a. ,, EDD) � 4CID) .. axm 0 +-'--,----,.......-.-.--......,....... --....... .-L--i D 2 4 6 Tme(tburs) 18 Figure I. Pulse labeling ofHaCaT cells with [14C)-Melanin. Total labeling in dpm per well at times 0, I, 2,4, 6, and 18 hours post label. Data is the average dpm/well of three wells and representative of three experiments. •p 0.05 compared to time zero. In addition, we have used l4C DOPA labeled melanin isolated from melanocyte for the pulse labeling of HaCaT keratinocytes and observed similar decreases in counts over time ( data not shown). However, natural melanin is also known to contain various proteins and these may stiU be protease sensitive sites. Since melanosomes are known to be engulfed by keralinocytes and partitioned into lysosomal bodies, we hypothesized that lysosomal inhibitors would suppress this degradation. Chloroquine, one such lysosomal inhibitor was observed to suppress the degradation and incorporation of synthetic melanin. This process requires new protein synthesis since cycloheximide was found to inhibit it. This further supports the theory that melanin is degraded in lysosomes

2003 ANNUAL SCIENTIFIC MEETING 'i 5000 .a .!! 4000 "o Dtotal label •= 3000 untreated i 'ti .. 2000 C CL ■total label c 1000 .!! +chloroquine GI E o o 1 2 18 time (hours) Figure 2. Effects of cyclohex:imide on radiolabeled melanin content in HaCaT cells. HaCaT cells were treated with 0, I, 2, and 4 ug/well of cyclohex:imide. Total counts in cells and media counted at time zero and 24 hours. Qata is the average dpm/well of three wells and representative of three experiments. Microsomal membrane prepared fractions from HaCaT cell cultures demonstrated the ability to solublize synthetic melanin. This enzyme appears to localize to the lysosomal fraction and have a pH optimum of pH 4 (data not shown). The protein nature of this enzyme is further supported by the demonstrated heat inactivates this extract. 5.5) 5.3) � 'ii' 5.10 ! 49J - • wtide � 470 s. 45) - - - 2%Kqi:add f 43) \. ,, I 410 --1% i 3.9J '.:�- ... . ., A!prgll.s 0dra:2 370 --l..l1r'!l!IB:I 35) ..... Dily7 0,,,9 ll,/12 Dily14 To test for the effects of a melanin degrading enzyme on skin color, we investigated the topically application of a fungal melanase. Aspergillus fumigatus e."tracts where applied to human skin that had previously been irradiated with UVB. On kojic acid, a known inhibitor oftyrosinase, and the Aspergillus extract both reduce enhanced the skin color loss after UVB. The above data supports the hypothesis that human skin degrades melanin and this process contributes to skin color. References: l.Ohtaki. N., Seiji, M. (1971) Degradation ofmelansomes by lysosomes. J. Investig. Dennalol. 57:1-S. 2. Saito, N., Seiji, M., (1976) Degradation ofmelanosomea in vitro and in vivo. Pigment Cell. 3:384-392. 117
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)