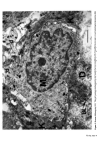
Causes of skin colouration 623 epidermal cells across these end caps. Some years later Cruickshank and Harcourt (7) were able to confirm this process of pigment donation in vitro in elegant cinematographic studies. Fitzpatrick and Breathnach (8) suggested the term 'Epidermal Melanin Unit' (Fig. 1) to describe a structural, as well as functional, use of the melanocyte and surround- ing keratinocytes. Its use should emphasize the fact that pigmentary processes involve not only the melanocyte but also the surrounding keratinocytes, which acquire the pigment secondarily. Fi•m 1. The epidermal melvin unit. Den•itic pro•sses of a basal •11 layer melanocyte (M) wind betw•n keratinocytes (K). Pigment granules, melanosomes, produced in •e melanocyte •e transferred to •e keratinocytes via the den•itic processes (•rows). BL: basal lamina. ORIGIN OF THE MELANOCYTE Experiments on embryonic mice led Rawles (7) to conclude that melanocytes arise from the neural crest (a strip of specialised ectoderm flanking each side of the neural Figure 2. Origin of the melanocyte. Migration (arrow) of embryonic neural crest cells (*) to the epidermis, mucous membranes, retina and central nervous system.

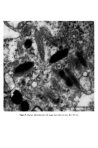
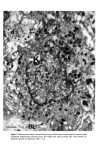
624 J. •1. •1. Hunter plate) and not from the neighbouring epidermal cells. Cells indistinguishable from mesenchymal cells, migrate from the neural crest and undergo differentiation into cells producing pigment when they reach the epidermis. This occurs as early as the fourteenth week in the human (Breathnach and Wyllie (10)). Once in the epidermis the melanocytes form a self-replicating population of cells, which retain their ability to migrate, even in adult epidermis. Conditions like neurofibromatosis, where pigmentary abnormalities are associated with nerve and suprarenal tumours, are experiments of nature which serve as a reminder of the pleuri-potentiality of neural crest cells. FINE STRUCTURE OF THE MELANOCYTE At the ultrastructural level the melanocyte is a distinctive cell. It is distinguished from neighbouring keratinocytes by its dendritic nature, its lack of tonofibrils, and des- mosomes are absent (Fig. 1). It is seen most often in the basal layer. Unlike the kera- tinocyte, mitochondria are abundant. Micro fibrils, as distinct from tonofilaments, are seen in the cytoplasm. Unlike tonofilaments they show no tendency to form bundles and are often seen as parallel arrays of fine filaments. The Golgi apparatus is usually prominent and the endoplasmic reticulum well developed (Fig. 3). The cell has a smooth plasma membrane which shows occasional thickened areas when apposed to the basal lamina. There are however no hemidesmosomal attachments to the basal lamina. The characteristic organelle of the melanocyte is the melanosome (Figs 3, 4, and 5). These are numerous in melanocytes of negroids and caucasoid individuals with a dark complexion though they are seen without difficulty in the melanocytes of those with a fair skin. They are ovoid or rod shaped bodies measuring 0-4-1.0 •tm in length and 0.1-0.5 I•m in diameter. Definition of melanosome (11): 'A melanosome is an organelle that is surrounded by a unit membrane and contains a highly organised internal structure of longitudinally orientated strands of concentric lamellae that have a regular pattern of dense particles with a characteristic periodicity. The organelles may be spherical or ellipsoid and often contain tyrosinase.' Drochmans (12) studied high resolution pictures of melanosomes and considered that the first stage in formation of the melanosome was the deposition of a protein matrix composed of coiled filaments (Fig. 4) Figure 4. Stage II melanosome (after Drochmans).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)