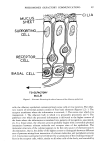
PHEROMONES (OLFACTORY COMMUNICATION) 53 performance changes with amounts of circulating estrogen and progesterone in the rat (35). Peak olfactory acuity occurs around the ovulatory period and the fluctuations are not evident in ovariectomized rats. A similar behavior is manifested in women. LeMagnen (36) found that the ability of women to detect the odor of a musk (Pentade- canolide) reached a maximum near or on the day of ovulation. Many materials of both animal and vegetable origin display a musk odor (37) and it has been often stated, with little substantiation, that musk odorants are of biological significance to man. Although the considerable variation in olfactory acuity with regard to Pentadecanolide was reported to be specific for this chemical (36), subsequent investigators (38,42) demonstrated increased sensitivity for at least eight different odorants in the time near ovulation. This cyclic variation correlates with alterations in hormonal levels occurring during the ovarian cycle. Men and ovariectomized women show no variation of olfac- tory acuity with time. Women with decreased gonadal secretion who were not receiv- ing estrogen often displayed poor odor sensitivity. Estrogen treatment significantly enhanced, but testosterone lowered, their acuity to odorants. Since menstruation is triggered by a decrease in estrogen levels, it is not unexpected that odor thresholds in women reach a maximum during the period of menstrual flow. The prepiriform cortex and preoptic area have particularly high uptake of estradiol, an estrogenic hormone, in the female rat (43). It is tempting to speculate that the increased estrogen liberated during woman's ovulatory phase of the menstrual cycle is accompanied by the hor- mone's increase in these regions of the olfactory pathway, thereby causing a heightened olfactory sensitivity to odorants. Despite our ignorance of the precise man- ner by which gonadal hormones influence odor sensitivity, a consistent finding in most studies is the well defined correlation between circulating level and type of hormone with the performance of the sense of smell in the female. In higher animals, the mechanism to ensure mating involves more than one sensory modality and often may require a complex interaction of diverse influence mediated by all the sensory processes. For example, women possess lower thresholds, i.e., greater sensitivities, to visual (44) and auditory (45) stimuli at approximately the time of ovula- tion. From the viewpoint of evolution, such peak attuning to sensory signals would increase the probability of coitus and result in an increased probability of conception. In man's dim past, increased sensitivity to an olfactory signal (acting as an aphrodisiac pheromone) continually emanating from the male might have been perceived by women near the time of ovulation only. Odor quality also may play a role in the olfac- tory behavioral response. The perceived quality of certain odors changes on dilution. Consequently, even if a biologically important odor is detectable throughout the menstrual cycle, the perceived odor quality may change with perceived intensity fluc- tuations. Until recently, anecdotal and indirect observations have indicated that social grouping of the human female can cause synchronization of menstrual timing. Such an effect is not unlike that seen in rodents (7). McClintock (46) conducted a study with young women living in the same college dormitory and demonstrated that menstrual syn- chrony indeed occurred. In addition, the data indicated a parallel with the Whitten ef- fect (8) in mice in which the suppression of estrus in groups of female mice is released by the introduction of a male pheromone. The suppression of the ovulatory cycle in some of the women was released when they were exposed to men. Odor may be impli- cated in these phenomena, but further studies must be conducted in order to de- termine the basis of the intriguing effects.

54 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS 4. LEARNED VERSUS INATE RESPONSES TO ODORS From Aristotle's proposal (47) that the various categories of animals might be arranged on a graded scale of complexity or perfection with man at the top, came the general ac- ceptance that all animals could be ranked on a single continuous dimension known as the scala nat•rae. However, such a "phylogenetic scale" is inconsistent with contempo- rary views of animal evolution. Hodos and Campbell (48) commented, "Comparative psychologists have failed to distinguish between data obtained from living representa- tives of a common evolutionary lineage and data from animals which represent di- vergent lineages. Only the former can provide a foundation for inferences about the phylogenetic development of behavior patterns." Primates evolved as a special branch of the insectivore line, e.g., shrews, moles and hedgehogs, dating back more than 60 million years. Over this period of time, carnivores and rodents have followed inde- pendent and different courses of development from the primate line and from each other. Consequently, from the aspect of the evolution of primate characteristics, com- parisons such as rat-cat-man are meaningless. The rodent olfactory system can be com- pared to man's because they are basically similar despite the fact that there is no phylogenetic relationship. A clue to patterns of evolution in the human lineage could be gained from a comparison among living insectivores, prosimians, Old World monkeys and man. It was for this reason that special importance was accorded to recent work (11-16) pertaining to the influence of vaginal odors of female rhesus monkeys on the sexual behavior of the male rhesus. The proposed existence of a primate phero- mone sparked interest in the search for human pheromones. Before discussing the putative rhesus monkey pheromone and its implication for hominids, let us briefly review some recent knowledge and thoughts relevant to learned responses to odor. The pheromone concept implies that learning and/or prior experience with odorants do not play an essential role in its effect. However, it is now generally recognized that the original definition of pheromone is misleading, even for insects. The olfactory sti- mulus is not always simple (49) and insect behavior may be more plastic or modifiable by previous experience than heretofore suspected (50). In other words, an insect's response to pheromones is not necessarily programmed by genetics. Since mammalian behavior is infinitely more complex and more flexible, responses to pheromones are intricate, variable and often experience-oriented (51). Because of the strong de- pendence on experience, Bronson (51) has proposed the term s/gna///ng rather than re- /eas/ng for those odors yielding an immediate effect on mammalian behavior. Mam- malian responses to sex pheromones are far more easily modified by early associations than has been supposed (52,53). Similar to avian imprinting, there often appears to be a sensitive period for olfactory imprinting (54). Early experiences with artificial odorants, ranging from cage shavings to arbitrarily selected perfumes, have modified sexual behavioral preferences, eating, aggression, exploration, etc., in lower animals (53). For example, weanling rat pups showed a preference for citral subsequent to in- jecting their lactating mothers with this chemical. Not unexpectedly, there is a paucity of information pertaining to the sense of smell in human neonates. Newborn infants respond to sensory stimuli and their suckling behavior changes on presentation of various odorants (55). Suckling ceased or decreased in frequency on exposure to iodoform, anise or peppermint oils in approximately the same percentage of children as was observed in rabbits, kittens and puppies. Russell (56) reported the existence of olfactory maternal attraction in a recent study on human neonares. Sleeping infants were presented with breast pads previously worn by breast-feeding mothers. Two-day
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)