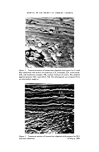
SEX ATTRACTANTS IN PRIMATES 267 Treatment c•38.•74. 6- E 0 Treatment 30 - g, 67. ? 76. 2O 50 40 30 20 I0 E -- -- _ E E E E Withdrawal I (a) , Withdrawal Ee (b) Donor receives progesterone (25 mg daily) •, (c) Male onosmic Nose unblocked Jd3B.•45. .•E (d) IOF2E 2E E 0 / I I I I I I I I I I • I I 2 q 6 8 I0 12 I• 16 18 20 22 2q 26 28 Days •i•re 7. Ch•n•es in the •exu•l peffotm•n• (mountin• p•ttem•) of m•le thesu• monkeys fo]lowin• odout •h•n• of their female •ttnet. the olfactory sense with an attractive female initially improves the sexual performance of the male, and briefly increases his ejaculatory score. It would appear from these results that at least two separate neural pathways are involved in the integration of olfactory cues. It is possible that sexual attraction involving releaser pheromones is brought about via olfactory connections to the pyriform cortex and other cortical regions

268 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS before passing on to the limbic system via the amygdala and medial fore- brain bundle to the hypothalamus. This I envisage as serving a filtering function, with the cortex analogous to a computer incorporating certain 'go' and 'no go' programmes. As we have already seen, the rhesus monkey's behaviour incorporates a number of'no go' programmes, as for example the modifying effects of partner preferences, past experiences with certain females, and the presence of other males, etc. If we consider the human with infinitely complex behaviour patterns involving traditions, rituals, religions and past experiences, there are many variables which might constitute 'no go' programmes, making it pointless to look for any overt behavioural response. If, on the other hand, we consider sexual performance (as in Fig. 7 for the rhesus monkeys) it is possible that a more direct neural input to the hypothalamus is involved, possibly monosynaptic or probably involving only a few synapses. This I see as being analogous to tactile input as in genital stimulation (1), with the hypothalamus serving as an area of integra- tion, while the threshold for response is modulated by hormonal status. In conclusion, olfactory cues are of importance in the sexual behaviour of the male rhesus monkey but the behavioural response is far from stereo- typed. This presents obvious problems when considering an attractant role for such odours in the human where social and cultural determinants of behaviour are even more complex. I would like to suggest that an alternative approach for the human might be in considering a role for these odours in sexual performance. This could be analysed objectively, and is less likely to be influenced by cultural variables. (Received: 31st December 1974) REFERENCES (1) Barraclough, C. A. and Cross, B. A. Unit activity in the hypothalamus of the cyclic female rat: effect of genital stimuli and progesterone. J. Endocrinol. 26 339 (1963). (2) Bertrand, M. The behavioural repertoire of the stump-tailed macaque. Bibliotheca Primat. 111 (1969). (3) Blurton-Jones, N. G. and Trollope, J. Social behaviour of stump-tailed macaques in captivity. Primates 9 365 (1968). (4) Carpenter, C. R. Sexual behaviour of free ranging rhesus monkeys (Macaca mulatta) I1 Periodicity of oestrous, homosexual, antoerotic and non-conformist behaviour. J. Comp. Psychol. 33 143 (1942). (5) Curtis, R. F., Ballantine, J. A., Kerverne, E. B., Bonsall, R. W. and Michael, R. P. Identi- fication of primate sexual pheromones and the properties of synthetic attractants. Nature 232 396 (1971). (6) van Hoof, J. A. R. Facial expressions in higher primates. Syrup. Zool. Soc., Lond. 8 97 (1962).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)