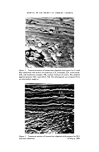
JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS '. .. • . z'•'• •'• . •.-..• ..... .,• • . :• -.... • • •:. •'.•--. •.• -r • ß • . , .. Figure 6. Transverse hair section digested with pronase for 100 h and then stained with PTA. Figure 7. As Fig. 5 but stained with ammoniacal silver nitrate. (.Facing p. 294)

CHEMISTRY OF HUMAN HAIR CUTICLE--III 295 In the light of the foregoing results we consider that the amino acid analysis of the material from whole cuticle which dissolves after 5 days treatment with pronase is representative of the proteins in the whole endocuticle. This analysis is designated ENDO in Table I. The endocuticle evidently contains more aspartic acid, glutamic acid, threonine, alanine, isoleucine, leucine, methionine, tyrosine, phenylalanine, histidine, lysine and arginine and less serine, proline, glycine, valine and cystine + cysteic acid than whole cuticle. This is in very good agreement with the analyses made by Bradbury and Ley (3) for the corresponding com- ponents isolated from wool cuticle with pronase (cf. columns 8 and 9 of Table I). By comparing the analysis for the endocuticle not only with that of whole cuticle but also with that of the exocuticle (cf. column 3 and the next section of the dis- cussion) it is evident that the major proportions of the acidic and basic amino acid residues in the cuticle are contained within the endocuticle. This is in accord with electron histochemical observations concerning the distribution of acidic and basic groups within the cuticle (1). That the amount of cystine in the endo- cuticle is quite small is also in agreement with electron histochemical observations (4, 5). (b) Exocuticle + A-layer + inner layer + cell membrane complex (EXO + A + I+ M) We have established that treatment of whole cuticle with pronase for 5 days dissolves the major portion of the endocuticle perhaps leaving some cystine-rich exocuticular-like material and cytoplasmic lipid debris in the endocuticle region. It follows that the material remaining after pronase digestion will comprise the inner-layer, exocuticle, A-layer and cuticle cell membrane complex. Bradbury and Ley (3) have isolated a similar fraction from wool cuticle with pronase and al- though the exocuticle will certainly comprise the major proportion of this fraction, A-layer and membrane components will also be present. Our analysis for the long term pronase-insoluble material is shown in column 3 of Table I. As would be expected, the analysis is complementary to that of the whole endocuticle in that it contains more serine, proline, glycine, cystine + cysteic acid and less aspartic acid, glutamic acid, threonine, alanine, valine, iso- leucine, leucine, methionine, tyrosine, phenylanaline, histidine, and arginine than whole cuticle. There is excellent agreement between these results and those of Bradbury and Ley (3) for the comparable exocuticle-rich fraction isolated from wool cuticle (cf. columns 8 and 10 of Table I). That the exocuticle-rich fraction contains high concentrations of cystine and lower concentrations of the acidic and basic amino acids relative to the endocuticle is also consistent with electron histochemical observations. Of particular interest is the very high cystine concentration in the long term pronase-insoluble fraction in which « cystine occupies 1 in 4 of the amino acids of
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)