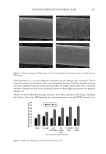
589 SUPPRESSION OF ITCHING BY THREE HERBAL ETHANOLIC EXTRACTS investigate whether the three medicinal herbs affect FLG mRNA expression, we treated HaCaT cells with IL4 + IL13 in the presence or absence of A houstonianum, B falcatum, S chinensis, and a combination of the three. We found that the downregulation of FLG mRNA expression induced by IL4 + IL13 was significantly (p 0.01) restored by single or combination treatment, as revealed by RT-PCR (Figure 3D) and qPCR (Figure 3E). The decrease in the abundance of pro-FLG protein by IL4 + IL13 stimulation was significantly (p 0.001) restored in the presence of A houstonianum, B falcatum, S chinensis, and a combination of the three (Figure 3F). Double immunofluorescence staining showed that IL4 + IL13 stimulation decreased the punctate patterns of FLG protein staining (Figure 4). However, pretreatment with a combination of the three extracts restored punctate staining similar to that of the control cells. These data suggest that A houstonianum, B falcatum, and S chinensis alone or in combination may help repair damaged skin barrier function by restoring reduced FLG expression in keratinocytes. THE THREE HERBAL EXTRACTS INHIBIT IL4–INDUCED IL31 EXPRESSION Th2 lymphocyte-derived cytokines (e.g., IL4 and IL31) transmit the itch sensation by directly activating sensory neurons and blocking IL4 and IL31 signaling by neutralizing antibodies reduces itch in patients with AD (1,44–46). IL31 is considered a crucial mediator of histamine-independent itching (47). Keratinocytes release multiple pruritogenic factors in response to Th2 cytokines (48). However, it is unclear whether keratinocytes express IL31. We tested whether IL4 stimulates IL31 mRNA expression in HaCaT keratinocytes. RT-PCR showed that IL4 enhanced IL31 mRNA expression in a time-dependent manner Figure 4. Effect of the three herbal extracts on FLG subcellular localization in HaCaT cells. HaCaT cells cultured on coverslips were treated with IL4 (20 ng/mL) plus IL13 (50 ng/mL) in the absence or presence of the combination of the three herbal extracts (40 μg/mL A houstonianum, 20 μg/mL B falcatum, and 40 μg/mL S chinensis) for 36 h. After fixing and permeabilization, the cells were incubated with anti-FLG primary, and Alexa Fluor 488-conjugated secondary antibodies (green fluorescence) for immunofluorescence staining. Rhodamine-labeled phalloidin was used as the counterstain for F-actin (red fluorescence). Nuclear DNA was counterstained with Hoechst 33258 (blue fluorescence). The areas in the dashed boxes are magnified in the right panels. Scale bars, 400 μm.

590 JOURNAL OF COSMETIC SCIENCE (Figure 5A), and qPCR revealed that the increase in IL31 mRNA levels was 1.72 ± 0.068-, 2.07 ± 0.208-, 3.03 ± 0.416-, and 1.67 ± 0.153-fold at 6, 12, 24, and 36 h of the control, respectively (Figure 5B). The increase in IL31 protein level was also increased in a time- dependent manner after IL4 stimulation (Figure 5C). In addition, ELISA demonstrated a time-dependent increase in IL31 protein levels secreted into the culture medium by IL4 stimulation (Figure 5D). After 24 h of IL4 stimulation, secreted levels of IL31 peaked, increasing from 23.4 ± 6.96 pg/mL (control) to 113 ± 7.51 pg/mL. These results imply that IL4 stimulates IL31 expression at the transcriptional level in HaCaT cells. We next investigated the effects of the three medicinal herbs on IL4-induced IL31 expression at the transcriptional and translational levels. RT-PCR (Figure 6A) and qPCR (Figure 6B) analyses showed that A houstonianum, B falcatum, and S chinensis significantly (p 0.05) inhibited IL4-induced IL31 mRNA expression in HaCaT cells. Furthermore, their combination treatment inhibited IL4-induced IL31 expression more effectively than each treatment. Similar to mRNA levels, IL4-induced accumulation of IL31 protein was decreased to control levels after treatment with the herbal ethanolic extracts (Figure 6C). As determined by ELISA, pretreatment with each herbal extract significantly (p 0.001) reduced IL31 concentration in the culture medium, and the combination treatment of the three extracts was more effective than their single treatment (Figure 6D). This was supported by the results of double immunofluorescence staining, which showed that IL4 stimulation increased IL31 staining in both the cytoplasm and nucleus. However, exposure to a combination of the three reduced the intensity of IL4-induced IL31 staining, like that of the vehicle control (Figure 7). Our findings on IL31 expression suggest that A houstonianum, B falcatum, or S chinensis inhibit IL4-induced IL31 production and that the combination of the three extracts has a better effect than each extract singly. Figure 5. IL4 upregulates IL31 expression in HaCaT cells (A and B). HaCaT cells were treated with IL4 (20 ng/mL) for various time periods (0–36 h). Total RNA was isolated, and IL31 mRNA levels were examined using RT-PCR (A) and qPCR (B). GAPDH was used as the loading control (C). HaCaT cells were treated as in (B), and whole-cell lysates were immunoblotted using anti-IL31 antibodies. GAPDH was used as the loading control (D). HaCaT cells were treated as in (B), and the concentration of IL31 in the culture medium was determined by ELISA. *p 0.05, **p 0.01, ***p 0.001 compared to control (n = 3).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)