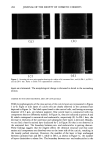
284 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS _3. Own (a) (b) (c) (d) Figure 1. Scanning electron micrographs showing the surface of (a) untreated hair, and (b) DS-1, (c) DS-2, and (d) DS-3 hair. Refer to Table I for experimental conditions. layers are eliminated. The morphological change is discussed in detail in the succeeding section. CHANGE IN THE CROSS-SECTIONAL AREA OF CUTICLE CELLS TEM microphotographs of the cross section of the cuticle layers are represented in Figure 3 (a-f). Eight or nine layers of cuticle cells are clearly observed in the untreated hair depicted in Figure 3a. The black spots found in the cortical cells, and having an average diameter of 0.3 Ixm, are melanin pigment. One cuticle layer is constructed with two components that are differently stained. In Figure 3a, they are indicated as points A and B, which correspond to exocuticle and endocuticle, respectively (9). In DS-1 hair, the decrease in thickness of the outermost and subsequent three layers is detected. Besides, we can find a heavily stained layer (indicated by C in Figure 3b) that is not observed in the untreated hair. The boundary between exo- and endocuticles is getting obscure. These findings suggest that the reagent affects the chemical structure and that some amino acid components are dissolved even on the inner side of the cuticle, resulting in the loosely packed structure. However, the number of the layer is kept unchanged between untreated hair and DS-1, while in DS-2, as shown in Figure 3c, the number of layers diminishes to about five. The boundary between exo- and endocuticles in the

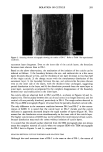
ISOLATION OF CUTICLE 285 Figure 2. Scanning electron micrograph showing the surface of DS-2'. Refer to Table I for experimental conditions. outermost layer disappears. Even on the inner side of the cuticle layers, the boundary becomes more obscure than in DS-1. Based on the above observations, the mechanism of the isolation of the cuticle can be inferred as follows: 1) the boundary between the exo- and endocuticles in a few outer layers becomes obscure at first, and the thickness of one layer decreases to less than half of the virgin cuticle 2) the change occurs with the simultaneous dissolution of the outermost layer 3) the boundary between the exo- and endocuticles becomes obscure even on the inner side of the layers, with the thickness kept unchanged, suggesting partial dissolution in the layer and 4) the dissolution proceeds from the outer layer to inner layer, successively accompanied by the complete disappearance of the boundary between exo- and endocuticles in the inner layer. No cuticle cells are observed both in DS-3 and DS-4, as shown in Figures 3d and 3e, respectively. The boundary between the cuticle and cortex is no longer detected, and the cortical cells are partially dissolved, particularly in DS-4. The rough surface detected in DS-3 by an SEM micrograph (Figure ld) arises from the partially dissolved cortical cells. The only difference in the treatment condition between DS-2 and DS-2' is the concen- tration of KOH. It is noted that the cuticle layer in DS-2' shrinks and the cuticle thickness is apparently decreased despite the equal number of layers (Figure 3f). This means that the dissolution occurs simultaneously on the inner side of the cuticle layer. The higher concentration of KOH may not be preferred for mild removal of hair cuticle, because dissolution may reach the cortex without isolation of cuticle layers. It is noted that the smooth surface observed from the SEM micrograph does not always mean the complete removal of cuticle when we compare SEM with TEM micrographs for DS-2 hair in Figures lc and 3c, respectively. RELATION BETWEEN CUTICLE THICKNESS AND REACTION TIME Although the total treatment time of DS-2 is the same as that of DS-l, the extent of
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)