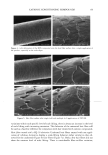
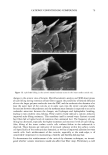
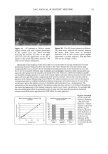
2002 ANNUAL SCIENTIFIC MEETING 89 Table I: Dielectric Constants of Common Lipophilic Sunscreen Ingredients (Ingredients listed in BOLDFACE t7pc were used in the study.) Chemical Isoeicosane Polyisobutene Mincral oil Octyldodecyl neopentanoate Ethylhcx¾1 stearate Eth¾1hcxyl palmirate Isopropyl palmirate lsopropyl myristate Ci 2-15 alk•l benzoates Caprylic/Capric tri$1¾cerides Dicth¾1hex¾1 adipate Dibutyloct•l malate But•loctyl salicylate Diethylhexyl malate Octyl methoxycinnamate Octyl salicylate Diisoamyl malate Avobenzone lsododecyl alcohol dimer with IPDI Dimethyl capramide Oxybenzonc Dielectric Constant •, 25øC 2.04 2.13 2.13 3.04 3.05 3.06 3.20 3.24 3.78 3.83 4.21 4.65 5.35 5.88 5.98 6.20 7.42 io (est.) 10 (est) i 2.43 i 3.0 (literature reference) Table II: Oil Phase Compositions, Measurements, and Results for Test Formulations Containin, 'late, 3% Ox•'benzone, 2% Avobenzone Ingredients A - B (5 D E F G H I Isopropyl myristate 10% C12-15 alkyl benzoates 10% 10% Diethylhexyl malate 10% 15¾o 10% 7.5% Saturated hydrocarbon 5% 5% 5% C20 neopentanoate 5% Diisoamyl realate 10% 9.25% Dibutyloctyl malate 5% 4% Dimethyl capram/de 1.75% 7.5% C12 dimer with IPDI 5% Dielectric constant 4.57 4.92 5.48 5.89 7.44 7.66 7.83 8.71 10.59 Photodecay rate constant -.095 -.078 -.062 -.055 -.028 -.027 -.024 -.015 -.032 In vivo SPF 17 20 21 25 i Turro, N.J., Modern Molecular Photochemist•7, Menlo Park, Benjam/n/Cummings, 1978, p. 329 2 Marcus, R.A.d. chem. Phys. 1956, 24, 966. Marcus, R.A. Annu. Rev. Phys. Chem. 1964, 15, 155.

90 JOURNAL OF COSMETIC SCIENCE FRONTIERS OF SCIENCE AWARD LECTURE SPONSORED BY COSMETICS AND TOILETRIES© T Cells, Skin Immunosurveillance, and the Human Immune Response Thomas S. Kupper, MD Harvard Medical School, Boston, MA The immune system is charged with protecting the host from infectious agents. Such agents are encountered at interfaces with the environment, and the skin is our largest of these interfaces. Immunity can be thought of as having two components--the Innate Immune System and the Acquired Immune System. Innate Immunity is phylogenetically ancient and elements of it can be found in insects and plants. In human skin, protein mediators such as defensins (which directly kill bacteria), cytokines, and complement are released when skin is injured. These events recruit leukocytes to the compromised integument and optimize host defense. The Innate Immune System, however, cannot "learn"-•each response is identical to the previous one, both qualitatively and quantitatively. In contrast, the Acquired Immune System can change and adapt, developing what is tantamount to "memory". A much more recent evolutionary development, Acquired Immunity developed about the same time as vertebrates in associated with longer life spans. Acquired Immunity is based upon cells with antigen receptors--B cells and T cells. The reshuffling of segmentss of genes during development generates billions of different antigen receptor sequences. For example, T cells that develop in the thymus express receptors composed of ct[3 or 78 chains, and each T cell has a T cell receptor with a unique sequence. Antibodies (B cell receptors) recognize tertiary, three-dimensional conformations of proteins. T cell receptors, on the other hand, recognize peptides of 8-20 amino acids bound to Major Histocompatibility Complex (MHC) molecules. Class I MHC molecules, such as HLA-A, B, and C bind peptides derived from the cytoplasm (both self and viral peptides) and present antigen to CD8+ T cells. Class II MHC molecules, such as HLA-DR and DQ bind peptides derived from proteins internalized from the extra cellular space (e.g., from bacteria or fungi), and present antigen to CD4+ T cells. HLA molecules are expressed at highest levels on dendritic cells, which also express costimulatory molecules that help maximally activate T cells. When a T cell's antigen receptor encounters a dendritic cell's HLA molecule binding the correct antigenic peptide, the T cell becomes activated by antigen. If this is the first time that the T cell h as seen its specific antigen, the T cell proliferates, and one cell gives rise to thousands of progeny. Some of these progeny survive for many years. This clonal expansion means that more antigen specific T cells are available to respond the next time such an antigen is encountered. If this antigen is from a pathogenic bacterium, then this immunologic memory serves host defense. With so •aaany different T cell receptors, how doex the i•nmune system ever get the right T cell to the right place st tb, e right time? NaYve T cells, which have never encountercd antigen, are not allowed to roam freely in the body. Rather, they recirculate between blood and lymph nodes, specialized organs that concentrate lymphatic drainage from large surface areas interfacing the environment. This recirculation is dependant on the expression of specialized cell surface molecules on both naive T cells and lymphatic vessels. Dendritic cells in skin, including epidermal Langerhans cells, internalize pathogens and "process" their proteins into peptides, which are expressed on MHC molecules. During this process, many of these dendritic cells migrate from skin to lymph node via afferent lymphatics. NaYve T cells are channeled through these lymph nodes, and thus the environment is effectively concentrated in an anatomic area in which naYve T cells are abundant. This is an efficient means of immunosurveillance of skin, performed in the draining lymph node. Another form of immunosurveillance depends upon the ability of certain T cells to exit blood vessels in skin. These T cells have special "skin homing" molecules expressed on their surface that
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)