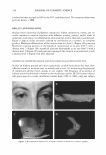
ADHESION FAILURE IN HAIR 353 = f Fiber Surface -------: r ! r !- � INNER BETA LAYER 008fOO_p_ep-ti-de-Hy_d_rog_ e_n---,-r- Hydrophilic Protein(�/ bonds&. Salt links DEL TA "Contact Zone" � Globular Protein (e)�OooJBoi LAY l ER Hydrophobic -Peptide & van der W aals fibrous protein (d) � - lB-MEA ( c� f f ---'------Van der W a :: ER B ET A L AYER Hydrophobic . __ _ I keratin protein �.l___J)isulfide �ioesterbonds & Peptide Keratin protein (a}-- bonds A LAYERS of CMC INTERLAYER BONDING Hydrophil�j_:i, �tide� H dr�g n- � protein f O O nD o Bonds & Salt Links : Globular Protein e-0ao O�eptide & : Hydrophobic � van der Waals 1 fibrous protein d f-� �� � -van der Waals : 18-MEA c-f f f"•u•L:: T _ -/-��A _ Th1oester ,, Hydrophobic � ,, ,, Keratin Protein b- nal.clx.il2nac: Disulfkfe � & Peptide Keratin Protein a- ' A-Layer--- ◄- - - - - - - -'PEROXIDE B Figure 2. A. The monolayer model for the cuticle-cuticle CMC, with principle bonding proposed between layers. The inner beta layer is the "lower" and the outer is the "upper" beta layer. B. The monolayer model for the cuticle-cuticle CMC (lower half) and its principle interlayer bonds, with some primary sites for chemical attack. two lipid layers (called inert beta layers) separated by intercellular cement, the delta layer (Figure 2A). Several important developments have occurred since Fraser's description of the CMC that have further improved our understanding of this important structure. Among these important developments are the work of Rivett and coworkers (3 ,4) and Bryson and coworkers (5 ). Two different models are presented in this paper in which this prior research is taken into consideration to provide a more detailed chemical represen tation of the CMC. In our models, we are proposing the interconnecting bonds between the different layers of the CMC to assist in our interpretation of hair-fracturing mecha nisms. Both of our models consist of protein and fatty acid layers. Model 1 (Figure 2A,B) contains monolayers in the beta layers, while model 2 (Figure 3) contains bilayers in the fatty acid domains consistent with Fraser's original concept. We are also suggesting a change in terminology. Up to this point, the beta layer immediately on top of each cuticle cell has been called the upper beta layer and the one beneath each cuticle cell has been called the lower beta layer. This terminology is confusing, especially for schematics of the CMC as in Figures 2A,B and 3. Therefore, we propose that the beta layer that sits on top of each cuticle cell and often becomes the outer hair surface be called the outer beta layer and the one immediately beneath each cuticle cell be called the inner beta layer (see Figures 2A,B and 3), and this terminology will used throughout this manuscript. Figure 2B shows only one-half of the CMC structure for the monolayer model and depicts some of the primary sites for chemical attack by permanent waves, bleaches, and sunlight. The top half of the CMC is not depicted in this figure because it is similar to the lower half, with the exception that it does not contain 18-MEA but contains saturated fatty acids (Cl2-Cl8) and oleic acid in place of 18-MEA. Our preference is for the monolayer model over the bilayer model for the following reasons: • Swift (2) has pointed out that a monolayer model fits better from the point of view of CMC measurements. • If the beta layers are monolayers, then 18-MEA is linked to the delta layer through short hydrophobic side chains rather than through ester or thioester bonds (in the

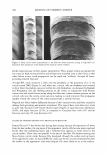
354 JOURNAL OF COSMETIC SCIENCE � Fatty acid--�--�� j I c 1 i 1 � r ...,..__INNER BETA LAYER Hydrophilic protei � f GI b I t 0 O OnOQo Peptide, Hydrogen DELTA o uar proe. � 0 OUos-,....- b d &Salt l· -d.J on s iks LAYER Hy�rophobic f brous t . 0 0009 0u /Peptide & van der Waals l 1 pro e1� □- 0 o-o.,:._...r Fatty aci � ____✓E_ .._ _e_ _ lin _ _a_g_ _ ______ _ 18-MEA----- I LJ +---OUTER BETA LAYER Hydrophobic } !�van der Waals keratinprotein----........._� } =.,..,,,hioester bonds Keratin protein �-Disul f ide & Peptide bonds Figure 3. The bilayer model for the cuticle-cuticle CMC, with principle bonding proposed between layers. bilayer model). Some of these hydrophobic amino-acid side chains in the monolayer model are also branched, making the outer beta layer even more susceptible to failure at the delta layer, where it has been shown to occur (7). • The monolayer model contains fewer layers of ester/thioester linkages thus it will be more specific in its reactions to nucleophilic agents like mercaptans, hydroxide, and hydroperoxide anions-a point addressed later in this paper. Another point of contention concerning the CMC is whether or not the delta layer contains globular proteins or glycoproteins (2). We believe the current evidence favors globular protein layers in the delta layer for the following reasons: • No further evidence for glycoprotein in the CMC has been presented since 1991 (8) however, Bryson et al. in 1995 (5) isolated lipid-soluble lipoproteins from the delta layer of cortex-cortex CMC and not glycoproteins, favoring the globular protein model. • The delta layer resists solubilization by reducing or oxidizing agents and by acids and alkalies (5). If the CMC contains globular proteins similar to those in many other membranes containing large domains of hydrophobic amino acids on their surfaces (9), then these should be resistant to aqueous reagents as Bryson found. Thus the globular protein model is consistent with its known reactivity. • The delta layer swells only about 10-15% (10) therefore, much of it is hydrophobic. Such a relatively hydrophobic delta layer is more consistent with a globular protein model than with a more hydrophilic glycoprotein.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)