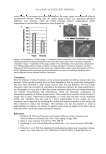
JOURNAL OF COSMETIC SCIENCE 476 Because cytoglobin appears to infl uence oxygen’s effects at the cellular levels, it has been suggested that cytoglobin may infl uence procollagen production by moving oxygen to key enzymes responsible for the early synthetic steps in collagen production (4,5,15). In particular, an enzyme called prolyl-4-hydroxylase, which is also a heme-based enzyme, requires oxygen to be attached at its active site in order to facilitate conversion of proline to hydroxyproline in early-stage procollagen production (Figure 1) (16). This paper will explore the possibility that plant-based substitutes for human globin proteins, namely leghemoglobin, may function in a similar capacity to assist procollagen production. All green plants express heme-based globin proteins, which are thought to function as free-radical scavengers and oxygen-transport molecules, much as they do in humans (17). While all green plants express globin proteins (called phytoglobins), cer- tain plants, namely legumes, which are known to fi x nitrogen, accumulate this protein in special structures found on the plant roots called symbisomes (generally known as root nodules). Pueraria lobata, a plant more commonly known as kudzu, is a legume that grows in many parts of the world. It grows rapidly and can extensively cover many other plants, which causes problem in countries where the plant has no real predators. The roots of kudzu contain symbisomes, and these can be easily harvested and extracted to provide useful quantities of leghemoglobin. The ability of plant-based hemoglobins to scavenge reactive oxygen species has been established (18). The paper will examine whether a plant-based extract containing leghemoglobin can infl uence the genes responsible for upregulation of extracellular matrix proteins in normal human dermal fi broblasts compa- rably to human-derived cytoglobin. MATERIALS AND METHODS PUERARIA LOBATA (KUDZU) SYMBIOSOME EXTRACT AND PUERARIN Using previously published methods, an aqueous extract taken from root nodules isolated from Puerraria lobata (kudzu) symbiosomes was standardized to 0.2–0.5 mg/ml leghemoglobin (3). Purifi ed puerarin was obtained from Sigma Chemical Company (Milwaukee, WI). Figure 1. The proposed role of cytoglobin to act as an oxygen shuttle to deliver oxygen to prolyl-4-hydrox- ylase, which, in turn, oxidizes proline residues to hydroxyproline residues in early-stage procollagen (steps a–b). The collagen fi gure was adapted from reference 16.

KUDZU EXTRACT AND PROCOLLAGEN PRODUCTION 477 MICROARRAY ANALYSIS—NORMAL HUMAN DERMAL FIBROBLAST CELL CULTURES Cultured normal human dermal fi broblasts (NHDF, Cascade Biologics) were grown in T-75 fl asks until confl uent using appropriate culture conditions. Upon reaching con- fl uence, the cells were treated with culture media supplemented with test materials at the concentrations specifi ed or with culture media alone, which acted as the untreated control. After applying the test materials to the cells, they were incubated for 24 hours at 37 ± 2°C and 5 ± 1% CO2. At the end of the incubation period, the culture media was re- moved by aspiration and the cells were washed once with cold phosphate-buffered saline using approximately 1 ml per well. After the washing step, a trypsin/EDTA solution was added to the wells to release the cells. After the cells were released, an appropriate vol- ume of trypsin-neutralizing solution was added to the wells. The treated cells and the untreated cells were pooled into separate 15-ml centrifuge tubes and pelleted by centri- fuging at 1000 rpm at 4 ± 2°C. After removing the supernatant, the pelleted cells were lysed by adding 300 μl of guanidinium thiocyanate lysis solution to each tube and then repeatedly drawing and releasing the solution into a pipet until the cell pellet dissolved. The cell lysate was then stored at −75°C until the RNA extraction process could be completed. RNA ISOLATION (AMBION RNAqueous KIT) To the thawed fi broblast lysate described above was added an equal volume of 64% etha- nol, and the culture tubes were mixed via vortex mixing. Seven hundred microliters of the alcoholic mixture was transferred to a glass-fi ber fi lter cartridge, the fi lter cartridge was loaded into a 1.5-ml collection tube, and the entire apparatus was centrifuged for 1 minute at 14,000 rpm. The fi ltration process was repeated until all of the mixture was fi ltered. The fi lter was then washed to remove any residual cellular debris from the RNA bound to the glass fi bers by subsequently applying 700 μl of wash solution followed by an additional two washes with 500 μl of wash solution, each time removing the wash solution by centrifuging the tube at 14,000 rpm for one minute. The fi ltrate was spun to remove any residual wash solvent. RNA bound to the glass fi bers within the cartridge was eluted by applying 30 μl of Tris-EDTA buffer (10 mM Tris-HCl, 1 mM EDTA, pre- heated to 70–80°C) to the cartridge and centrifuging the cartridge in a new collection tube at 14,000 rpm for one minute. After the RNA had eluted, it was analyzed as de- scribed below. RNA CONCENTRATION ASSAY (Molecular Probes Ribogreen Assay) Ribogreen reagent was provided as a stock solution in DMSO. Prior to use, the reagent was diluted 2000-fold in TE buffer. The RNA assay requires 200 μl of diluted Ribogreen reagent per sample to be tested and 1 ml of the reagent for standards. Once prepared, the diluted reagent was stored in the dark. A series of RNA standards was prepared by diluting purifi ed ribosomal RNA derived from E. coli to the following concentrations: 2 μg/ml, 1 μg/ml, 200 ng/ml, 40 ng/ml, and 0 ng/ml (blank). Prior to assaying, the RNA samples, prepared as described above, were diluted 1000-fold in TE buffer. For the RNA assay, 100 μl of the diluted samples or standards
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)