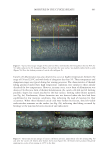
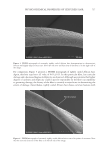
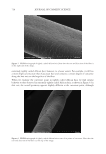
645 WHAT CAUSES CURLY HAIR? data have become the basis of standard measurements in both the wool and hair industries. In the wool industry, automated microscopy techniques (notably the optical fiber diameter analyzer) rely on measuring curvature from small subsamples of single fibers from within multifiber “staples” (a tress) and generating curvature in units of degrees per millimeter (15). In hair research, the routine approach is to constrain single hairs under a transparent sheet and line up hairs against a circular ruler to give a radius or diameter in millimeters or centimeters (Figure 1C). Loussouarn et al. (16) developed a curl classification system based on hybrid measurements where curl diameter was augmented by other simple two- dimensional measurements that indicate elements of the three-dimensional shape of a hair, including along-hair variability. Combining measurements in a statistical model allowed origin-independent classification of hair curl into a system of type I (straightest) to type VIII (most three-dimensionally curly). DIFFERENT TYPES OF CAUSATION AND NONCAUSATIVE CORRELATION The question “what causes and influences hair curl” is more complicated than it first appears. Causation occurs in multiple contexts. Curvature and torsion in a mature, dead, hair shaft are caused by the chemistry and structural organization of the shaft. That shaft structure was caused by processes during hair growth in the follicle, including the spatiotemporally distributed processes of keratin maturation and cornification. Ultimately, germline genes and physiological factors defined the follicle’s program to grow a hair of specific curliness, with this cause being first implemented during embryogenesis and then reconfigured by successive cycles of telogen and anagen. What causes curliness at the level of inherited genes may be of little interest to on- the-head product developers, but the cause of curliness at the level of the hair shaft itself might inform new technology. Conversely, dermatologists, anthropologists, and forensic scientists will differ in what level of causality is relevant. Irrespective of whether we are investigating curliness at the level of the hair shaft, hair growth, or follicle cycle, causation occurs within a complex system. A hair shaft contains multiple layers (i.e., medulla, cortex, and cuticle) and sublayers (e.g., orthocortex, paracortex) that were derived from once-living cells (17). Within the cortex, for example, hair shaft structural organization can be described over a wide range of spatial scales, with each having potential contributions to fiber structure and mechanical properties. These include protein chemistry (e.g., the matrix and the keratin intermediate filaments it surrounds), the architecture of filaments into macrofibrils, and the overall fiber shape. Within an anagen follicle, the situation is more complex again because the follicle is a miniature organ that creates a tissue that then cornifies into a range of different hair morphologies. Mammalian hair diversity indicates that a wide range of functional fibers can be produced by a follicle. While investigating causes of curliness, we should therefore be mindful of the following: • Curliness may have multiple potential causes (e.g., fiber ellipticity, cortical organization). • The multiple causes may be nonexclusive therefore, a specific curliness is an emergent phenomenon (i.e., the sum of causes). • There may be features that correlate with curl, but do not cause curl, that are caused by a common third phenomenon.

646 JOURNAL OF COSMETIC SCIENCE As researchers have come to realize that hair shaft properties may be emergent rather than due to a single cause, it has been recognized since the early 2000s in wool (18) and more recently in hair (19,20) that this requires investigation using a systems biology approach. Within this complex system, we expect to see some phenomena that correlate with curliness but do not cause curliness. This is a common problem in biology called a spurious correlation. A version of the issue was popularized by Gould and Lewontin (21) in the context of evolutionary biology. In the context of fiber growth, for example, various studies find that both the expression of inner root sheath keratins (22) and hair shaft ellipticity correlate with hair curl (23,24). But, does the keratin expression cause the ellipticity, or is it a consequence of ellipticity because follicles have a circular cross-section, and the inner root sheath fills in the difference between ellipse and circle? Or are both caused by a common causative mechanism, such as the organization of the neck of the dermal papilla that orchestrates the fate of cells into the various lineages including shaft and inner root sheath during hair growth (25)? This particular issue was first raised by Priestley (26), and despite decades of additional data slowly filling in our understanding of follicle biology, we still do not know the answer to this example question. An experimental approach is essential to directly tease out noncausative correlations from causation in addition to comparative analysis. WHAT FIBER FEATURES CORRELATE WITH CURL? WITHIN THE MATURE HAIR SHAFT Internal organization of the hair cortex, follicle morphology, the shape of the fiber cross- section, and differences in protein abundance have been described in the literature as features that correlate with curl. For more than 90 years, researchers have been trying to work out how fiber curvature correlation with mammalian hair features might explain a mechanism causing curliness. Studies have largely focused on three contexts: human scalp hair, midside wool from sheep, and mice (typically as models of genetic disorders affecting human scalp hair). Although there are some common features that correlate with hair shaft curl in scalp hair, sheep wool, and mouse hair (plus other species), the differences are particularly informative for identifying and discounting features underpinning hair curliness in all mammals. For example, studies of human scalp hair often point out that curly hair shafts tend to have a more elliptical cross-section than straight hairs (27–29). Ellipticity alone is not a mechanism, but could an elliptical profile be immutably connected to the process that generates curl? Sheep wool indicates that this is not the case, and there are clear examples of the reverse situation where hair shafts from merino and merino relatives that have a more elliptical profile are less curly than those with circular profiles (30). This demonstrates the challenge of correlating just one feature with curl. It has been appreciated since the 1950s that cell-type distribution in the cortex region of the fiber correlates with various measures of hair curliness (31). Sheep wool with lower- diameter hairs (15–25 μm) is the most curly and has been investigated as a model to study fiber curvature because it is made of three distinct modes of keratin organization (ortho-, meso-, and paracortex) embodied within macrofibrils composed of keratin intermediate filaments and matrix (32,33). The macrofibril-containing cells of the hair shaft, once living in the follicle, now dead and transformed to structural elements in the mature hair, are
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)