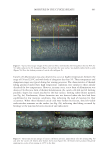
689 MOISTURE IN THE CUTICLE SHEATH of light interference that they produce. In the literature, it has already been shown that colorful patterns of light interference are produced when cuticle cells delaminate and buckle, creating micron-size air gaps that lead to the conditions for light interference (4–6). RESULTS AND DISCUSSION CUTICLE CELL LIFTING OR BUCKLING AT HIGH ELONGATIONS In the past, it has been shown that cyclical elongations with deformation values higher than 12% and in the moisture range between 40% and 65% relative humidity (RH) lead to patterns of cuticle cell buckling, forming parabolic shapes (3). Fig. 1a shows a hair fiber displaying cuticle cell buckling after being subjected to similar conditions. Unlike other types of cuticle cell buckling, which will be reviewed later, this type of damage is not caused by transient losses of moisture but rather by high elongations. Its formation, however, depends strongly on the equilibrium moisture content of the cuticle cells. The mechanism is as follows: when hair fibers are equilibrated at moisture conditions between 30% and 65% RH, and then subjected to elongations ranging between 5% and 17%, mechanical energy appears across the hair fiber in the form of tensile, shear, and Poisson stresses. In this range of deformation, the keratin intermediate filaments in the cortex undergo an alpha to beta transformation (7). This protein phase transition enables the cortex to unfold and relax, thereby allowing for higher elongations while partially dissipating high concentrations of mechanical stresses. In contrast, for similar elongations and humidity conditions, the cuticle sheath does not have such a phase transition mechanism to dissipate stresses. Thus, while the cortex can undergo high extensions without building up high levels of stresses, the cuticle sheath elongates in parallel but develops stress concentrations across its layers. The result is lifting and buckling of cuticle cells, as shown in Fig. 1a. Furthermore, Fig. 1b shows a magnified version of the fiber shown in Fig. 1a. In this picture, it can be seen that when cuticle cells lift under these conditions, there are no endocuticular remnants left on the cuticle cell Figure 1. SEM pictures of a hair fiber showing cuticle cell lifting and buckling at different magnifications Fig. 1a (×500), Fig. 2 (×3000). Lifting was produced by applying 10 cycles of 15% elongation at 50% RH.

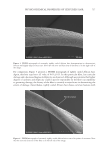
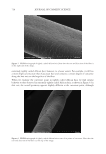
690 JOURNAL OF COSMETIC SCIENCE immediately below. This observation suggests that because of its multilayer composition, the top cuticle cells resist shear and tensile stresses produced by elongation. However, they fail at the CMC under the action of hoop or Poisson contraction stresses (3), which force them to lift upward. Cuticle cell failure by lifting and buckling is, thus, how the composite structure of the cuticle sheath dissipates stresses at high elongations. Cement breakage occurs at the CMC because at such elongations and moisture conditions, the CMC is still rigid and does not allow a compliant transfer of hoop stresses to neighboring cuticle cells underneath. By the same token, this analysis also indicates that the moduli of epicuticles, exocuticles, and endocuticles are stress hoop compliant, and, therefore, there is no damage across their boundaries. Yet the mechanism of stress dissipation changes from cuticle cell lifting or buckling to transversal crack formation and cuticle sheath detachment when the hair fibers are equilibrated at higher moisture contents (see Figs. 2a, 2b, 3a, and 3b). For instance, as the equilibrium moisture content increases between 65% and 80% RH, the number and area size of buckling patterns decreases, while the number of transversal Figure 2. Optical Microscopy (2a and 2b) and SEM (2c and 2d) pictures of hair fibers after subjected to 10 cycles of 15% elongation at 75% RH. Note the appearance of deep transversal cracks accompanied by small area size cuticle cell buckling patterns in all four pictures.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)