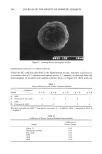
178 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS cent product 4-MU, as well as a concentration range for which the fluorescence inten- sity of this species varies linearly with concentration. A solution of 4-MU, 2 •g/ml, was prepared in 0.1 M Tris buffer, pH 7.0. The absorption spectrum was scanned, giving a maximum at 334 nm. The emission maximum, 444 nm, was obtained by scanning the emission spectrum while irradiating at 334 nm. All quantitative fluo- rescence studies employed these parameters. Linear calibration curves (Figure 4) were generated for the concentration ranges, (1) 0.2 •g/ml-2.0 •g/ml and (2) 0.02 •g/ml-0.2 •g/ml. The levels of 4-MU generated in beta-G and AS assays fall into range 1. The conversion factor was 1 fluorescence unit = 0.09 •M 4-MU. Samples were prepared as described above. All components (except the enzyme) were warmed to 37øC before addition of the enzyme. During the assays, reaction mixtures were maintained at 37øC. Fluorescence intensities were measured as a function of time. ODOR PRODUCTION IN APOCRINE SECRETION Sterile, odorless apocrine secretion was eluted from capillaries into 1.5 ml of 0.1 M Tris buffer, pH 7.0. In the first study, apocrine sweat was treated with beta-glucuronidase 12 10 o o.o ! 4-MU concentration, ug/ml Figure 4. Fluorescence as a function of 4-methylumbelliferol concentration. 0.2

AXILLARY MALODOR 179 (E. coli) and aryl sulfatase (Aerobacter aerogenes) at 0.01 mg/ml in Tris buffer, 0.1 M, pH 7.0. The apocrine secretion was also treated with the lipophilic diphtheroid and with a mixed culture of axillary bacteria, each in sterile saline suspension. In the second study, inhibitors of the two enzymes were included in the reaction mix- tures--Zn + + as ZnGlycinate, saccharic acid lactone, and hexametaphosphate. Samples were prepared in gas chromatography vials and sealed with crimp caps. Samples were incubated at 37øC. In the first study, odor evaluations were conducted at 16 hr and 40 hr of incubation. In the second study, odor evaluations were conducted at 24 hr and 48 hr. Odor evaluations were performed by a panel of five, including one professional perfumer. ENZYME ACTIVITY IN CELL-FREE MEDIA Lipophilic diphtheroid was grown for 18 hr in brain-heart infusion containing 0.1% Tween 80, centrifuged for 20 min, and the supernatant medium filtered through a 0.22-micron filter. The substrate solution (2 ml of 4-MUG or 4-MUS) was mixed with 1 ml of the filtered medium (or water, as control). RESULTS PILOT STUDY A double-blind screening study was designed to detect the enzymes in axillary secre- tions. On plates designed to test for beta-G, ten sweat samples gave high fluorescence nine of these came from "high-odor formers." Only one came from a "low-odor former." On plates designed to test for AS, six of eight "high-odor formers" were found to have swabbings containing AS (two samples were lost), and only two of the ten "low-odor formers" had the enzyme. This study shows that at least some axillary bac- teria generate steroid esterases and that the presence of these enzymes may be correlated with axillary odor. SEMIQUANTITATIVE ASSAYS OF ENZYMES: EFFECTS OF INHIBITORS Various inhibitors of mammalian beta-Gs and ASs have been reported in the literature (18-32). We tested several of these compounds as well as related materials in our assay. The classic inhibitor of beta-G is saccharic acid-A-lactone (glucarolactone) (24). We tested this material as well as the simple sugars, mannose, fucose, and galactosamine, and the polysaccharide pullulan. One of the reported inhibitors of beta-G, EDTA (21), is a strong chelating agent. Thus we also tested o-phenanthroline, citrate, gluconate, nitrilotriacetate, and hexametaphosphate against the enzyme. Table I summarizes the materials tested. Several materials were found to inhibit each enzyme, but three of the agents tested-- Zn + +, Cu + +, and hexametaphosphate--inhibited both beta-G and AS. QUANTITATIVE ASSAYS OF BETA-G AND AS The effects of five inhibitors of beta-G and three inhibitors of AS identified in semi-
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)