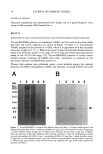
HAIR DAMAGE WITH S100A3 ELUTION 21 80 .c 60 • 40 o 20 (^) lOOO 800 600 400 200 0 1.5 800 T (c] • • 600 '-1 E E 400 0.5 .- o +.,e 200 o o (D) Root Middle Tip Root Middle Tip Figure 4. S100A3 and protein contents in the effluents and extractions of natural hair. Root, middle, and tip ends were subjected to an elution test under reducing conditions and to total protein extraction (See Materials and Methods). The amounts of S100A3 and protein were estimated through western blot analysis and protein assay, respectively. Elution of S100A3 (A) and protein (B) from the three parts of hair fiber under reducing conditions, and the contents of S100A3 (C) and protein (D) in the three parts, are presented. Each value is an average of eight samples. Bars indicate standard deviation. Significant differences in the values of the root-end part are indicated. *, p 0.05 **, p 0.01 ***, p 0.001. DISCUSSION Based on an earlier study that included fractionation of hair proteins on tricine/SDS/ PAGE (17), we have developed a procedure for sensitive and quantitative analysis of S100A3 derived from hair fiber. Application of this method to the extracts and effluents from hair fiber resulted in efficient separation of their constituent proteins (Figure 1A). A novel antibody was also prepared to identify the carboxyl terminal region of S100A3, which does not show any sequence homology with other members of the S100 protein family. On the basis of the recently proposed model for the three-dimensional structure of S100A3 (18), the carboxyl terminal region of S100A3 is likely to be the flexible part, which is not involved in dimer formation. Through western blot analysis of S 100A3, we were able to determine the accurate contents of S100A3 in both extracts and effluents derived from hair fiber (Figure lB). Furthermore, application of slot-blot analyses al- lowed an even more sensitive quantification of S100A3. It is known that the cuticular scales after prolonged waving treatment become delami- nated as evidenced by scanning electron microscopy. Since the endocuticle was identified as the weakest element of the cuticle structure subject to extensive damage especially during waving, it has been hypothesized that a loss of substances from the endocuticle can continue to hair damage (16). This is further supported by our recent studies employing ultrastructural immunocytochemistry, which showed S100A3 incorporation in the inner part of endocuticle (7).

22 JOURNAL OF COSMETIC SCIENCE We have explored whether S100A3 is stable following exposure to permanent waving lotion. The protein could react with reductants or oxidants and might be involved with the breaking and reforming of disulfide bridges in hair proteins (19). Cysteine residues of S100A3 are thought to be involved in cross-linking reactions via disulfide bonds. In particular, it is important to clarify the stability of the S100A3-rich layer (i.e., the inner part of the endocuticle). Our result showed substantial amounts S100A3 in the effluents of permanent waving lotions (Figure 2). It may be concluded from this result that the scale lifting resulting from the permanent treatment may be a consequence of degra- dation of the S100A3-rich layer. Delaminated cuticle was also frequently observed with hair fiber that had been exposed to repetitive shampooing and drying treatments (20). These treatments resulted in a release of material from the endocuticle (21). This kind of hair damage can be also associated with weathering, including exposure to UV radiation (22). It has been pro- posed that the amounts of eluted protein correlated with the degree of hair damage (23,24). Also, we were able to show that S100A3 was eluted from UV-irradiated hair, as well as permed hair, under non-reducing conditions (Figure 3). A possible mechanism of damage may include cleavage of the disulfide bridges of S100A3 by UV radiation. Our result suggests that the combination of daily UV irradiation and shampooing may contribute to a gradual release of S100A3 from hair and ultimately result in cuticle delamination. A contrast profile was obtained between elution of S100A3 and protein from distinct parts of hair fiber under reducing conditions. The amount of eluted S100A3 was lower in the tip end of hair as compared to the root end (Figure 4A). In contrast to this, the total amount of protein was higher in the tip end of hair (Figure 4B). This result is consistent with previous data showing that greater amounts of protein were extracted from damaged hair than from intact hair (23,24). In addition, the data suggest that almost all of the S100A3 to be released has already been lost in the tip section of hair. Thus, the tip part has a lower proportion of S100A3 than total extractable protein (Figure 4C,D). The difference between the proportion of S100A3 to protein in the effluents (15% from the root end, 1.6% from the middle, and 0.4% from the tip end [compare Figure 4A versus 4B]) may reflect that S100A3 is preferentially eluted from hair during everyday grooming operations. Therefore, S100A3 is affected to a larger extent by weathering as compared to other proteinous hair structure constituents. This susceptibility of S100A3 may be attributed to its localization in cuticle and to its solubility in water. The results of our experiments suggest that S100A3 is solubilized and eluted in the course of hair damage. Given the fragility of the S100A3-rich layer, we propose a new four-stage model that assumes S100A3 elution is a major cause of hair damage (Fig- ure 5): STAGE I Newly emerging hair from a follicle is characterized by smooth-edged scales. STAGE II A mechanical stress in intact hair can lead to cracking of the endocuticle (25). As a result
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)