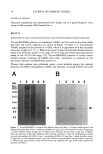
HAIR DAMAGE WITH S100A3 ELUTION 23 Intact cuticle (I) Grooming Perming Actual cuticle (•) I UV-exposure . S100A3 -'• ........... ' ' - Daily Damaged (lifted) cuticle washing Denatured cuticle (•v) (•) e Cross-I inked S100A3 e Denatured S100A3 Figure 5. A model of hair damage emphasizing the loss of S100A3 from cuticle. Smooth-edged scales are present in the hair shaft emerging from the scalp (I), but cuticle can be chipped away at the edges of the scales through normal grooming (II). Environmental stresses, namely UV radiation, affect the disulfide bonds or conformation of the S100A3 molecule located in the bottom part of the endocuticle (III). S100A3 is gradually eluted in daily washings of hair, which leads to the delamination of cuticle (IV). Treatment with permanent waving lotion alone can lead to the elution of S100A3 without environmental stresses. of normal grooming, cuticle fragments can be chipped away (26). The S100A3-rich layer becomes the outermost layer of hair readily exposed to hair treatments. STAGE III Environmental factors, including UV-radiation, can affect the disulfide bonds or con- formation of the S100A3 molecule. At this stage, the cross-links of cysteine residues of S100A3 are cleaved, and S100A3 becomes a soluble protein that differs from other insoluble hair constituents. Therefore, this chemical process would cause latent but considerable damage to the S100A3-rich layer. STAGE IV During daily washing, S100A3 would be gradually released from the hair that had been exposed to environmental stresses. Application of permanent waving lotions can elute S100A3 from the natural hair of stage II without undergoing stage III transformation, because the disulfide bridges are not resistant to the reductant and oxidant. Loss of S100A3 from endocuticle could result in scale lifting.

24 JOURNAL OF COSMETIC SCIENCE CONCLUSIONS Causes of hair damage, such as permanent waving, UV-irradiation, and weathering, all affected the binding of the S100A3 molecule to the adjacent cuticles. As a consequence of this, S 100A3 can be preferentially eluted from the inner part of the endocuticle during hair damage leading to the lifting of cuticular scales. S100A3 appears to be an important molecule closely associated with hair damage. REFERENCES (1) L.J. Wolfram and M. O. Lindemann, Some observations on the hair cuticle,J. Soc. Cosmet. Chem., 22, 839-850 (1971). (2) J. G. Gould and R. L. Sneath, Electron microscopy-image analysis: Quantification of ultrastructural changes in hair fiber cross sections as a result of cosmetic treatment,J. Soc. Cosmet. Chem., 36, 53-59 (1985). (3) K. Kizawa, H. Uchiwa, and U. Murakami, Highly expressed S100A3, a calcium binding protein, in human hair cuticle, Biochim. Biophys. Acta, 1312, 94-98 (1996). (4) B. Moore, A soluble protein characteristic of the nervous system, Biochem. Biophys. Res. Commun., 19, 739-744 (1965). (5) D. Engelkamp, B.W. Schiifer, M.G. Mattei, P. Erne, and C.W. Heizmann, Six S100 genes are clustered on human chromosome lq21: Identification of two genes coding for the two previously unreported calcium-binding proteins S100D and S100E, Proc. Natl. Acad. Sci. U.S.A., 90, 6547-6551 (1993). (6) K. Kizawa, S. Tsuchimoto, K. Hashimoto, and H. Uchiwa, Gene expression of mouse S100A3, a cysteine-rich calcium binding protein, in developing hair follicle, J. Invest. Dermatol., 111,879-886 (1998). (7) T. Takizawa, T. Takizawa, S. Atai, K. Kizawa, H. Uchiwa, I. Sasaki, and T. Inoue, Ultrastructural localization of S100A3, a cysteine-rich, calcium binding protein, in human scalp hair shafts revealed by rapid-freezing immunocytochemistry, J. Histochem. Cytochem., 47, 525-532 (1999). (8) U. G. F/Shr, C. W. Heizmann, D. Engelkamp, B. W. Schiifer, and J. A. Cox, Purification and cation binding properties of the recombinant human S100 calcium-binding protein A3, an EF-hand motif protein with high affinity for zinc, J. Bid. Chem., 270, 21056-21061 (1995). (9) M. M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding, Anal. Biochem., 72, 248-254 (1976). (10) H. Schiigger and G. von Jagow, Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa, Anal. Biochem., 166, 368-379 (1989). (11) J. A. Swift and B. Bews, The chemistry of human hair cuticle. I: A new method for the physical isolation of cuticle,J. Soc. Cosmet. Chem., 24, 13-22 (1974). (12) B.C. Powell and G.E. Rogers, "The Role of Keratin Proteins and Their Genes in the Growth, Structure and Properties of Hair," in Formation and Structure of Human Hair, P. Joll•s, H. Zhan, and H. H/Scker, Eds. (Briskhiiuser Verlag, Basel 1997), pp. 59-148. (13) S. Kanetaka, K. Miyata, and Y. Nakamura, Characterization of nonkeratinous proteins extracted from human hair by permanent wave lotion, J. Soc. Cos. Chem. Jpn., 24, 5-12 (1987). (14) M. Oku, H. Nishimura, and H. Kanehisa, Dissolution of proteins from hair. II: The analysis of proteins dissolved into permanent waving agent and the evaluation of hair damage,J. Soc. Cos. Chem. Jpn., 21, 204-209 (1987). (! 5) S.S. Sandhu and C. R. Robbins, A sensitive fluorescence technique using dansyl chloride to assess hair damage, J. Soc. Cosmet. Chem., 40, 287-296 (1989). (16) G. Mahrle, W. Sterry, and C. E. Orfanos, "The Use of Scanning-Electron Microscopy to Assess Damage of Hair," in Hair Research, C. E. Orfanos, W. Montagna, and G. Stuttgen, Eds. (Springer-Verlag, New York, 1981), pp. 524-528. (17) R. Kon, A. Nakamura, N. Hirabayashi, and K. Takeuchi, Analysis of the damaged components of permed hair using biochemical technique, J. Soc. Cosmet. Chem., 49, 13-22 (1998). (18) G. Fritz, C.W. Heizmann, and P.M. H. Kroneck, Probing the structure of the human Ca 2+ and
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)