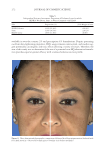
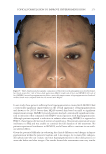
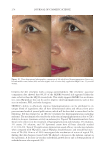
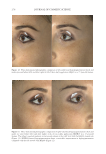
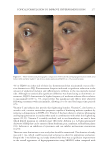
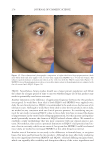
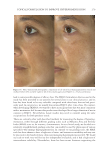
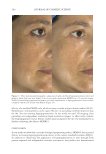
JOURNAL OF COSMETIC SCIENCE 324 Alzheimer’s disease, diabetes type II, and skin aging. Chronic exposure to intrinsic and extrinsic factors by itself generates the release of infl ammatory mediators, causing infl am- mation as well as dermal and extracellular matrix damage (2). The next section highlights the main pathways of skin infl ammaging, considering DNA damage and oxidative stress as key players in the process. OXIDATIVE STRESS Oxidative stress, which occurs because of a mismatch between ROS production and the cell’s ability to detoxify these species (30), is a key contributor to skin aging (31). ROS are generated by-products of oxygen metabolisms in every cell from different sources, including mitochondria, peroxisomal activity, oxidase activity, and endoplasmic reticu- lum (ER) (32). ROS at normal levels have benefi cial functions for the body, including cellular structure synthesis, fi ghting pathogens, and numerous signaling pathways (30). However, if ROS levels increase, oxidative stress occurs, harming cellular structures and immune responses and accelerating skin aging (30). Under normal conditions, receptor protein tyrosine phosphatases (RPTPs) inhibit recep- tor tyrosine kinase (RTK) activity on the cell surface through dephosphorylation (33). However, under oxidative stress, ROS bind to cysteine of RPTPs, inhibiting its activity and increasing phosphorylated RTKs levels. Consequently, numerous downstream signal- ing pathways are triggered, such as initiation of mitogen-activated protein kinase, tran- scription factor activator protein-1 (AP-1), and nuclear factor-κB (NF-κB) (34). The process inhibits collagen production and increases MMP gene transcription (2,35). Col- lagen degradation by MMPs leads to a build-up of fragmented and disorganized collagen fi brils, which downregulate new collagen synthesis (25). MMP-1, MMP-3, and MMP-9 collectively account for most of the type I and III dermal collagen degradation (36). In addition, reduced collagen content is attributable to AP-1’s suppression of type I and III procollagen gene expression in the dermis (37). D ifferent studies have reported a strong correlation between oxidative stress and infl am- mation, as continued exposure to ROS induces cell damage and, subsequently, a proin- fl ammatory signaling response. Oxidative damage to cells triggers TNF-α release, which in turn binds to cell surface receptors, activating the NF-κB infl ammasome. NF-κB in- fl ammasome generation results in an overproduction of other proinfl ammatory cytokines, which can be detrimental to health (38,39). In addition, UV radiation activates infl am- matory mediators, such as neutrophils, to remove damaged cells. Moreover, macrophages infi ltrate the exposed area, release ROS, and degrade the extracellular matrix (2,35). This is accompanied by fi broblast deterioration and an inability to repair the extracellular matrix, leading to skin aging (40). Therefore, NF-κB infl ammasome is considered the major etiology of infl ammaging. MICRO INFLAMMATORY THEORY The m icroinfl ammatory theory describes skin aging as a number of events in a repeated cycle that occurs because of cell exposure to intrinsic or extrinsic factors. (i) Damaged cells secrete proinfl ammatory signals such as prostaglandins and leukotrienes. (ii). These

SKIN-AGING AND INFLAMMAGING TREATMENT 325 signals bind to mast cells, causing TNF-α and histamine release. (iii) TNF-α and hista- mine stimulate intercellular adhesion molecule-1 (ICAM-1) synthesis in endothelial cells of the blood vessels. (iv) ICAM-1 binds to circulating immune cells that release hydrogen peroxide to perform diapedesis and enter the dermis. (v) Immune cells in the dermis re- lease singlet oxygen and MMPs, causing connective tissue damage. (vi) Immune cells reach the damaged cell, release hydrogen peroxide, and digest the damaged cell. (vii) During the three oxidative bursts, other cells are damaged, proinfl ammatory mediators are release, and the infl ammatory cycle is maintained (11). The mic roinfl ammatory theory could explain a number of skin aging features such as loss of elasticity, and dermal fl exibility. Moreover, this theory accounts for wrinkle appearance and epidermal thinning with age (12). Bhattacharyya et al. (13) assessed the histological changes in intrinsic aged mice. Results showed that there was a notable epidermal thin- ning and reduction in the pilosebaceous unit associated with aging (13). DNA DAM AGE UV radi ation creates pyrimidine dimers, resulting in DNA mutations. Moreover, UV radiation generates ROS, which accelerate telomere shortening and interfere with en- zymes required for DNA repair (41). Continu ous DNA damage responses cause replicative cell senescence and aging processes, as illustrated by Xia et al. (10). DNA damage in aging cells contributes to a surge in the proinfl ammatory secretory phenotype, which in turn induces further DNA damage and proinfl ammatory secretion in adjacent cells. Ultimately, local infl ammation becomes sys- temic, resulting in an infl ammaging exacerbation (10). Telomer es refer to nucleotide sequences at the ends of chromosomes that protect them from degrading (42). With each cell division, telomeres shorten because of replication problems. Accordingly, the proliferation capacity of the cell deteriorates, and eventually, cellular aging occurs (2,43). However, telomerase is an enzyme that retains telomere length by adding telomere repetitions to the end (44). Therefore, regulation of telomerase activity is a key factor in DNA repair and antiaging therapy. Goyarts et al. (45) conducted a study to investigate the molecular mechanism associated with morphological changes in age spots. mRNA in skin biopsies was analyzed, and re- sults revealed that 23 genes were upregulated, whereas 17 genes were downregulated. For instance, peptidase gene and genes responsible for keratinization and basement mem- brane synthesis were downregulated. Moreover, genes related to infl ammation were up- regulated, including MMP3, which is responsible for elastic fi ber degradation in the dermis. Both upregulated and downregulated genes were highly linked to infl ammation, confi rming the role of microinfl ammation in age spots (45). OBESITY Obesity , especially central obesity, is closely correlated with a proinfl ammatory state (39). This state is attributable to declines in subcutaneous adipose tissue and increases in visceral adipose tissue (VAT), which produces more infl ammatory cytokines with age (46,47). Elevated quantities of visceral fats are accompanied by higher levels of infl ammatory
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)