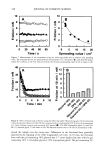
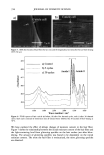
396 JOURNAL OF COSMETIC SCIENCE melanin: black-to-brown eumelanin and yellow-to-red pheomelanin (2). Eumelanin is believed to be a polymer derived from oxidative copolymerization of 5,6-dihydroxyin- dole (DHI) and 5,6-dihydroxyindole-2-carboxylic acid (DHICA) (3,4). Pheomelanin is composed of tyrosine and cysteine-derived units constructed into benzothiazine mono- mers that make the polymer (5). Extensive research work has been conducted by various groups to elucidate the structure of melanin and the melanogenesis pathway. Despite the vast literature on natural and synthetic melanins, the molecular structure of this class of pigments remains unknown. This is due to the lack of adequate methods to isolate melanin from biological sources, their insolubility at neutral pH, and the heterogeneity in their structural features. Melanins are typically isolated under harsh hydrolytic con- ditions such as prolonged treatment with concentrated hydrochloric acid or sodium hydroxide. Recent studies demonstrate that melanin, when exposed to boiling mineral acids, suffers profound structural alterations, particularly extensive decarboxylation (3,6). The isolation of melanin from hair or fur is more complex because of the com- pactness of the keratin in which the pigment is encapsulated. As a result, most of the literature work used Sepia melanin or synthetic eumelanin as a model for black hair melanin. Regardless of the source and type of melanin, various physical and chemical methods have been established in the last fifty years towards characterizing the structure of eumelanin and pheomelanin at the microscopic and macroscopic levels. Both X-ray and mass spectroscopic studies gained insight into the structural units of eumelanin and suggest that synthetic eumelanins and natural eumelanin from Sepia have similar build- ing blocks (7-9). Based on wide-angle X-ray diffraction measurements of eumelanin, a fundamental particle consisting of a graphite-layered structure with four to eight con- nected monomers per layer has been proposed. Both MALDI-mass spectroscopy and X-ray diffraction studies identified oligomeric units of masses in the range of-500- 1000 amu for Sepia eumelanin and synthetic eumelanin (8-10). Scanning electron mi- croscopy studies of size- selected Sepia melanin samples suggest that the pigment of eumelanin is an aggregated structure with subunits of-150 nm, and that smaller -15-nm-size particles adhere to these larger subunits (11). Recent AFM studies on Sepia melanin also confirmed that the 100-200-nm-size spherical particles are not a funda- mental structural unit of eumelanin, but are composed of smaller 5-15-nm-size particles (12). A review by Prota (13) claims the color of black hair is due to intact or poorly degraded eumelanin, whereas brown hair contains a degraded variant of eumelanin. Prota also claims that only red hair types contain pheomelanins. The study by Borgers et al. (14) uses sensitive HPLC methods for the identification and quantification of eumelanin and pheomelanin from human hair by measuring product yields produced by alkaline hy- drogen peroxide degradation of melanin. This study suggests that color determination for black, brown, or blonde hair depends more on melanin quantity than on eu-/ pheomelanin composition, while red hair may arise through production of pheomelanin. Wolfram and Albrecht (15) studied the oxidation of human hair with and without melanin pigment and the oxidation of melanin granules isolated from human hair. They found that hair with pigment degrades hydrogen peroxide at a much faster rate than hair without the pigment. Thus, they conclude that peroxide reacts with melanin much faster than with proteins. In another study, the same authors compared the chemical and photobleaching of brown and red hair and concluded that pheomelanin is more resistant

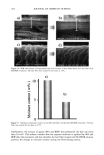
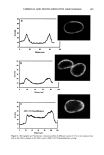
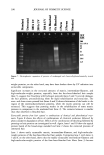
AMMONIA/PEROXIDE BLEACHING OF HAIR 397 than eumelanin to chemical or photodegradation (16). It is also known from the litera- ture that the ammonia/hydrogen peroxide combination is an effective melanin-bleaching agent, whereas hydrogen peroxide at neutral conditions bleaches melanin very slowly. Studies by Gallas et al. (17), using scanning tunneling microscopy (STM), showed a dramatic decrease in the STM-measured apparent heights for mildly bleached melanin compared to untreated melanin. These authors interpret the observed decrease as a result of oxidative conversion of the multilayered stacked sheets of melanin to largely destacked melanin sheets. There are no detailed studies in the literature on the ultrastructural features of hair meianosomes or on the mechanism of hair melanin bleaching induced by ammonia and peroxide. In order to study the mechanism of human hair melanin bleaching, intact hair melanosomes were isolated using an enzymatic procedure (1) to digest hair keratin under neutral conditions. The present study, aimed at understanding the mechanism of hair melanin bleaching, presents the first report on the direct demonstration by TEM of morphological and structural changes undergone by isolated hair melanosomes as a result of ammonia and ammonia/peroxide treatments. The study reveals that ammonia has a specific role in the bleaching process by breaking the melanosomal membrane, unlike other alkalizing agents such as sodium hydroxide and sodium carbonate at identical pH conditions. In addition, we also show by TEM that the micron-sized hair melanosomes are composed of smaller constituents of melanin (30-60 nm) encapsulated in a membrane sac. EXPERIMENTAL MATERIALS Untreated Asian black hair was purchased from International Hair Importers and Prod- ucts Inc., Bellerose, NY. Sepia melanin and all protease enzymes were purchased from Sigma Biochemicals (St. Louis, MO). All other chemicals and reagents used in the study were analytical grade. All solutions were prepared using milli-Q-water, and melanin suspensions were prepared freshly by sonication prior to the experiments. METHODS Isolation of hair eume/anin from Asian Mack hair. The isolation of black hair melanin was achieved by the procedure reported by Novellino et aL (1), using the following enzymatic treatments: (1) An Asian black hair sample, after removal of sebum and surface lipids, was treated with DTT (dithiothreitol) for 18 hr at pH 7.4 and 37øC. After centrifuga- tion, the pellet was treated with Proteinase K and DTT and stirred under argon at 37øC for another 18 hr. The mixture was centrifuged for 20 min (3500 x g, 4øC) in an Eppendorf centrifuge. (2) The pellet was extensively rinsed with water, suspended in 30 ml of 0.1 M phosphate buffer, and treated with papain and DTT at 37øC under argon for 18 hr. The mixture was centrifuged as above. (3) The black pellet collected after sixfold washing with water was suspended in 30 ml of buffer and treated with protease and DTT at 37øC under argon for 18 h. (4) The dark residue obtained after centrifu- gation was suspended in 40 ml of phosphate buffer deaerated with argon for 30 min. An oxygen-free solution of 2% Triton X-I00 was added and stirred at room temperature for 4 hr under argon. The mixture was centrifuged for 20 min (106,000 x g, 4øC). After
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)