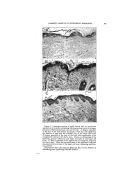
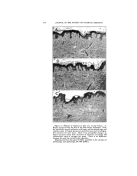
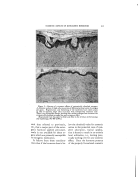
DETERMINING BACTERIOSTATIC POTENCY OF CHEMICALS 399 In offering this equation as a graphic exhibition of how bacterio- .: stasis may be brought about, and ::What effects it may eventually pro- duce, bsth on or in the cell as well as in the substrate, we make use of •the following assumptions. First, that the reaction between the bac- teria (B) and the chemical or medi- cation (M) is a simple stoichiometric one of the monomolecular reaction type, wherein one molecule of the •.. chemical reacts with one molecule :, of the cell constituents. We also assume that M is always in excess, .:•:. as would be the case if we have com- plete bacteriostatic or germicidal action. Furthermore we assume, as is actually present in use, that there exists in the medication- bacteria complex some organisms which do not get in contact with the medication because of such physico- chemical factors as film formation, agglutination, adsorption, clumping, etc., and thus do not react the same as far as killing or bacteriostasis is concerned, at least during the same time qnterval. Such organisms are represented as © and thus appear on both sides of the equation. The sum of the amounts of B on the right side of this equation must, naturally, equal 100 per cent of that which entered the reaction at the start (represented on the left). In just what proportion the total B is divided varies as described above. For the purpose of our discussion here we assume that little or none of the organisms exist in the spore form (M.B or true bacteriostasis), but that in the presence of high dilu- tions of certain chemicals the great majority of the cells will be as repre- sented by M.B. These eventually with time may become IM.BI and finally • when the cell dies (Chart 2, curve 3b). Or, eventually the cell by its own repair mecha- nism may neutralize the M and re- pair the damage done by the M.B q- IM.BI combination, acting over the extended period of time. Thus, M.A is formed and B is set free, ready to start reproducing again. (Chart 2, curve 3a). This explains the misnomered "Recovery from death" phenome- non and shows how it is brought about. That is, if during the time interval between where M.B and [ M.B ] are present, and before •--• is reached there is added an anti- dotal substance A to the mixture, it will bring about the reaction M.B q- [M.B Iq- 2A = 2B q- 2M.A (eq. 2, Chart 4). Thus the cell will eventually become free to grow once more. Practical conditions as represented here have occurred in the recent past in connection with the use of mercury compounds. These were formerly thought to be highly germicidal, but now have been found actually to be highly bacteriostatic and not germicidal in anywhere near the previously reported concentrations. In this same connection, so as to make this picture more complete, we wish to make a brief mention of the "Shippen technique," used for many years as a means for recog-

4OO JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS nizing both the existence and the extent of any bacteriostatic ififlu- ence on germicidal test data as de- termined by the standard phenol- coefficient procedure. In general, the method comprises making a subtransfer (4 loopfuls) from the original subtransfer tube. By this sub-subtransfer it is assumed that the toxic material M will be affected through dilution so as to make it no longer capable of acting bacterio- statically. That is M.B =M+B Time does not permit going further here to show certain fallacies in this procedure. Suffice it for our cur- rent purposes merely to mention the fact. Another phase of this b•cterio- static influence on micro-organisms, especially as to how it may cause false interpretations of germicidal test data, was shown by the speaker several years ago. Experimental data were presented then (1) which show how, even though a "-, 0" re- sult is obtained by the standard F.D.A. phenol-coefficient and the Cade-Halvorson plate count modi- fication tests, actually at the end- point time of the test (e.g., at the 10-minute contact time period) many viable organisms are present. These organisms may be considered to be, for the most part, in th'e IM.B[ condition and therefore die off dur- ing the incubation period. How- ever, some of them at least are capable of being neutralized under the influence of a proper antidote and the IM.B.[ = M q- B, setting free the bacteria to grow again. In other words, from the test find- ings it has been assumed that all of the organisms were dead--in real- ity they are not--but are actually in a condition of growing under suitable influences, i.e., they are still potential pathogens. Mention only can be made here, for lack of time, of one more set of conditions which exist in connection with the practical use of bacterio- static substances, as well as an inter- fering influence producing false germicidal test data. This includes the effects, especially, of such sub- stances as quaternary ammonium compounds, wherein some 99+ per cent of the test organisms apparently are truly killed in a short interval of time (even less than 1 minute), but a few bacteria remain viable (even for 30 minutes or more). By the standard F.D.A. phenol-co- efficient procedure a "-" result is always obtained by, say a 1-.q000 level, whereas by using such special techniques as the Klarmann-Wright semimicro method, (2), the Stuart ring or carrier method (3), the Cade swab technique ({), or others, it can be sh9wn , beyond any doubt, that some viable bacteria are still present. The reason for this situa- tion has been attributed by some to the fact that the quaternary compounds possess high bacterio- static properties and, therefore, pro-. duce these results in a manner simi- lar to that described above. That is: 2M q- 2B = M.B + and (M.B q-IM.B[ + 2A 2M.A + 2B).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)