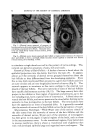
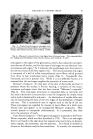
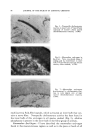
86 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS RELATION OF STRUCTURE TO FUNCTION It is possible to classify the end-organs that have been described accord- ing to the sensation they serve. Such a classification is found in Table 2. In all regions of skin the sensation of pain is served by the networks of dermal nerves. On the other hand, the end-organs that supply the sensa- tion of touch are quite varied, although almost all tactile nervous tissue is found about the hairs as baskets of nerve fibers. They range from complex end-organs, such as the networks of nerves of the hair follicles, to the muco- cutaneous end-organs. All known forms of nerve endings can serve the sen- sation of touch. This includes the network ofdermal nerves, which may per- ceive a rudimentary form of tactile sense in regions such as the cornea and, in general, in simple animal forms which have no other endings. Since all TABLE 2--DERMAL NERVE ENDINGS: CLASSIFICATION ACCORDING TO FUNCTION Type of Skin -Nerve Structure for Perception of Pain and Temperature Touch Haired skin Dermal network Hair-follicle network Glabrous skin General Dermal network Distal Dermal network Mucocut•neous Dermal network Mucous membrane Dermal network Complex papillary nerve Meissner's corpuscles and hederiform ending mucocutaneous end-organ Dermal network sensations may be perceived by one ending, the network of dermal nerves, and one sensation may be subserved by all the specialized endings developed in all the animal forms studied, it becomes apparent that the actual structure of the nerve ending is not of great importance. What is impor- tant is the particular cell of origin of a given cutaneous nerve fiber as well as the rami that lead its electrical activity into the tracts of the spinal cord and higher centers. In man, separation of the sensation of touch from the sensations of pain and temperature is effected at the level of the spinal cord and brain, but in lower animals, whose receptors are simple, no such segrega- tion is to be found. This once more emphasizes the fact that in the organ- ized central nervous system, there has not always been a grouping of nerve fibers according to sensations. Viewed from the phylogenetic heights, all structure and organization have evolved for the purpose of perceptual acute- ness, not for the purpose of providing special receptors for specific sensa.- tions. Anatomic data indicate that a pattern of sensation is presented to the central nervous system from the skin. Regarding the rabbit ear, Weddell and colleagues (4) have shown that approximately 100,000 hairs are sup- plied by 6000 neurons and that no regular distribution of the nerve fibers occurs. Many fibers are stimulated at a given point and a pattern of

THE SKIN AS A SENSORY ORGAN 87 sensation is produced. It is assumed that the spatial and temporal rela- tionships between nerve fibers and the conduction of impulses along them allow for the localization of a stimulus. In studies of conduction it has been shown that a rough grouping of fibers occurs: rapidly conducting touch fibers and more slowly conducting pain-temperature fibers (15). At the level of the spinal cord a fairly definite division of the pathways for touch and pain-temperature sensations exists. The dorsal tract is the newest, phylogenetically, and provides for specialized touch sensation. Wall and Cronly-Dillon (16) have recently described a region of receptor cells in the dorsal horn of the cat's spinal cord which they think may receive all stimuli and determine what is perceived and with what intensity. It is possible in the cat and in man to interfere with all experimentally induced sensations by producing a vibratory stimulus which "jams" the receptor-cell system. Thus, while the presence of a pattern of perception is indicated in the skin, its central connections may be through specific pathways or through a single neuron pool that filters out those perceptions that are of interest. At present we know little of the central connections and must feel fortunate to be able to recognize the pattern imposed by the anatomy of the peripheral nervous system of the skin. SENSATIONS IN ABNORMAL SKIN Itching is the most common pathologic sensation it accompanies most inflammation of the skin. In neurodermatitis, exquisite itchiness may be the principal symptom. It would be natural to assume that in an itchy skin there are more nerves than usual, but, as I have shown (5), there are fewer nerve fibers. It appears that chronic inflammation actually destroys nerve fibers. It is such destruction of patterns of nerve fibers that Weddell and associates (17) indicated was the cause of hyperalgesia. Repetition of a stimulus usually lowers the threshold of response thus, repeated itching could increase the reactivity of the local tissue to the sensation. Such reactivity could also be physiologically conditioned as a result of chronic inflammatory change. The presence of proteolytic enzymes in inflamma- tory fluids is well known Shelley and Arthur (18) have shown that enzymes such as trypsin and papain may induce local cutaneous pruritus. Generalized pruritus may also occur on such a basis. Section of the nerve leading to the skin will remove sensory capacity. While the complicated end-organ observed in the silver preparation de- generates under such circumstances, the histochemical reaction associated with the end-organ is maintained. In cats, dogs and monkeys, it xvas demonstrated that the histochemical reaction was still present one to four months after section of the nerve. Hurley and Koelle (19) exposed the skin to diisopropylfiuorophosphate by iontophoresis and were unable to change sensation. These two experimental findings suggest that the cholinesterase
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)