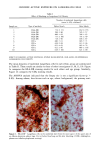
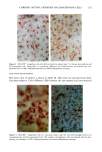
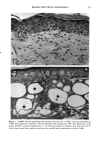
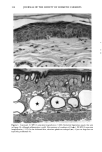
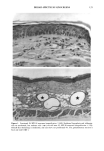
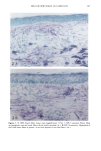
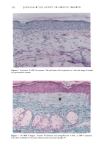
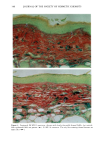
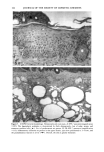
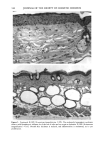
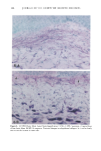
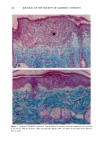
BROAD-SPECTRUM SUNSCREENS 153 ative of UVA-induced damage were the dense GAG deposits at the dermal-epidermal junction, especially prominent at 16 MPD (10). With stains for general morphology (H&E) and elastic fibers, it was not possible to distinguish between UVB and UVA effects. It is likely that transmitted radiation from both broad wave bands contributed to the observed damage. Effects of suberythemal UV doses have also been reported in human studies. Our observation of sunburn cells (SBCs) in most specimens at both dose points, regardless of the SPF of the sunscreen, confirms findings by Kaidbey in acute studies (16). Human subjects, pretreated with an SPF-15 sunscreen, were exposed to an SSR dose equal to the previously determined SPF of the product on each individual. Subjects were also treated with an SPF-30 sunscreen and irradiated with 15 MEDs. Faint erythema developed in some of the SPF-15 protected sites but not in the SPF-30 sites. Although the greatest number of SBCs occurred in the SPF-15 protected group, SBCs were induced in all subjects and with both sunscreens. Thus, injury to epidermal cells can occur in the absence of erythema and with UV doses that are far below the SPF of the sunscreen. In a different approach, but nevertheless demonstrating the effects of suberythemal radiation, Lavker et al. (17) exposed human subjects, at different sites, to 0.5 MED of SSR (290-400 nm) and 0.5 MED of UVA (320-400 nm) over a period of 5 ¾2 weeks for 28 doses. Notable changes compared to unirradiated skin were increased binding of lysozyme to elastic fibers, which suggests some change in the fibers, and increased inflammatory cells as visualized by leukocyte common antigen deposition. Similar changes were produced by both spectra, with the greatest effect seen with UVA. These studies, including our present findings, attest to the fact that structural components of skin can be damaged with far less radiation than it takes to produce a sunburn. In contrast to the evidence from our study, it has been proposed, from theoretical considerations, that the amount of UVA radiation transmitted through high-SPF sun- screens is not a significant factor in chronic photodamage (18). This a, gument derives from calculations based on acute exposure with erythema as the end point without considering the diverse biologic consequences of chronic exposure over decades. •', also fails to take into account the low absorbance capacity of some UVA II absorbers and the long-term effects of UVA on dermal matrix damage. For a UVA protection factor to be such that it is capable mainly of preventing an MED of UVA-induced erythema during the exposure period is not supported by in vivo experiments that show skin damage at suberythemal UVA doses. The claim that skin damage from 150 minutes of protected exposure with an SPF-15 sunscreen would be the same as from ten minutes of unpro- tected exposure (18) is therefore true only with respect to erythema. Recent research has demonstrated that although UVB is the most damaging portion of the sunlight reaching earth, UVA is responsible for numerous molecular and biochem- ical events that contribute to photodamage in skin (19-21). There is little dispute that both UVB and UVA contribute to chronic photodamage. Additionally, the effects of long-term UVA radiation are qualitatively and quantitatively quite different from those of UVB (10,22,23). Thus, protection against both broad wave bands is needed. The concept that prevention of sunburn is sufficient needs to be reexamined in light of the multiple effects demonstrated in this study of both UVB and UVA in the absence of erythema. Moreover, as our data show, extending protection well into the UVA I region with the addition of avobenzone to the octyl methoxycinnamate-oxybenzone sunscreen

154 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS considerably reduced all aspects of photoaging that were examined. Importantly, dam- age progression over time, from 10 MPD to 16 MPD, was especially moderated in the SPF-18 treated mice. These data support the concept that broad-spectrum protection should include UVA I absorption and that SPFs higher than 15 to 30 are needed to moderate the accumulation of photodamage over the many decades of human sun exposure. ACKNOWLEDGMENTS We wish to acknowledge the expert contributions of Marilyn J. Crosby for animal treatment, Dorothy Campbell for histologic preparations, and William Witmer for photography. REFERENCES (1) D. S. Snyder and M. May, Ability of PABA to protect mammalian skin from ultraviolet light-induced skin tumors and actinic damage, J. Invest. Dermatol., 65, 543-546 (1975). (2) L. H. Kligman, F. J. Akin, and A.M. Kligman, Sunscreens prevent ultraviolet photocarcinogenesis, J. Am. Acad, Dermatol., 3, 30-35 (1980). (3) L. H. Kligman, F. J. Akin, and A.M. Kligman, Prevention of ultraviolet damage to the dermis of hairless mice by sunscreens, J. Invest. Dermatol., 78, 181-189 (1982). (4) D. L. Bissett, D. P. Hannon, and T. V. Orr, An animal model of solar-aged skin: Histologic, physical and visible changes in UV-irradiated mouse skin, Photochem. Photobid. 46, 367-378 (1987). (5) S. R. Plastow, J. H. Harrison, and A. R. Young, Early changes in dermal collagen of mice exposed to chronic UVB irradiation and the effects of a UVB sunscreen, J. Invest. Dermatol. 91, 590-592 (1988). (6) L. H. Kligman, F. J. Akin, and A. M. Kligman, Sunscreens promote repair of ultraviolet radiation- induced dermal damage, J. Invest. Dermatol., 81, 98-101 (1983). (7) J. A. Harrison, S. L. Walter, S. R. Plastow, M.D. Bart, and A. R. Young, Sunscreens with low sunprotection factor inhibit ultraviolet A and B photoaging in the skin of the hairless mouse, Photodermatol. Photoimmunol. Photochem., 8, 12-20, (1991). (8) A. Fourtanier, J. Labat-Robert, P. Kern, C. Berrebi, A.M. Gracia, and B. Boyer, In vivo evaluation of photoprotection against chronic ultraviolet-A irradiation by a new sunscreen Mexoryl © SX, Pho- tochem. Photobiol., 55, 549-560 (1992). (9) L. H. Kligman and P. Zheng, The protective effect of a broad-spectrum sunscreen against chronic UVA radiation in hairless mice: A histologic and ultrastructural assessment, J. Soc. Cosmet. Chem., 45, 21-33 (1994). (10) L. H. Kligman, F. J. Akin, and A.M. Kligman, The contributions of UVA and UVB to connective tissue damage in hairless mice, J. Invest. Dermatol., 84, 272-276 (1985). (11) L. H. Kligman and R. M. Sayre, An action spectrum for ultraviolet induced elastosis in hairless mice: Quantification of elastosis by image analysis, Photochem. Photobiol. 53, 237-242 (1991). (12) Food and Drug Administration, Proposed monograph for sunscreens, Fed. Reg., 43, 38206-38269 (1978). (13) L. H. Kligman, Luna's technique: A beautiful stain for elastin, Am. J. Dermatopath., 3, 199-201 (1981). (14) R. H. Lesnik, L. H. Kligman, and A.M. Kligman, Agents that cause enlargement of sebaceous glands in hairless mice. II. Ultraviolet radiation, Arch. Dermatol. Res , 284, 106-108 (1992). (15) L. H. Kligman, Topical retinoic acid reduces tumorigenesis in hairless mice previously exposed to solar simulating radiation, Photochem. Photobid., 61S, 22S (abstract)(1995). (16) K. H. Kaidbey, The photoprotective potential of the new superpotent sunscreens, J. Am. Acad. Dermatol., 22, 449-452 (1990). (17) R. M. Lavker, G. F. Gerberick, D. Veres, C. J. Irwin, and K. H. Kaidbey, Ct mulative effects from
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)