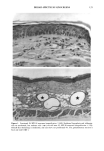
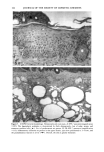
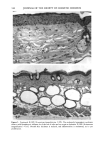
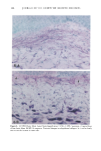
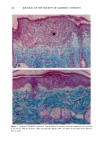
164 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS o pO.05 ' ß 3O 2O 10 0 Vehicle-Treated Y2-Treated Figure 5. Effect of natural lipid mixture on skin capacitance in normal human skin (n -- 20): 40 Ixl of Y2 (1.6%) or vehicle was topically applied to normal human skin (about 20 cm 2 area). Skin capacitance was measured before and two hours after Y2 or vehicle application. The data are expressed as percentage increase after Y2 or vehicle application. Results are mean - SEM. also accelerates barrier recovery after barrier disruption in both human and mouse skin. In contrast to physiologic lipids, nonphysiologic lipids, such as petrolatum, form a nonmembrane domain in stratum corneum, thereby improving barrier function (9). Furthermore, we showed that topical nonphysiologic lipids, such as petrolatum, induce earlier barrier recovery in comparison with physiologic lipids. Thus, it is more likely that the enhancement of barrier recovery by Y2 is due to acceleration of the formation of membrane bilayer structures in the stratum corneum rather than to the formation of a nonbilayer, hydrophobic domain as induced by petrolatum (9). However, more work needs to be performed to confirm this hypothesis. Moreover, further studies are needed to determine whether natural lipids complement or duplicate the properties of other inert species often used in cosmetics, e.g., glycerin, lanolin, and dimethicone. Never- theless, that the results of our human studies parallel our results in hairless mice further validates the application of the murine assay for assessing the effects of exogenous lipid mixtures on stratum corneum barrier homeostasis. The Y2 mixture primarily contains complex lipid precursors of the three key lipids, rather than the final products. It has been documented that there are large amounts of hydrolase activity in the upper epidermis (20-23). Moreover, we showed previously that triglyceride could substitute for free fatty acids, and glycosylceramides and sphingo- myelin for ceramides in the acetone model (8,13). Thus, we hypothesize that the basis for the efficacy of these complex lipids can be attributed to abundant enzyme activity in the upper epidermis, which would insure that these complex lipids will be catabolized rapidly to their nonpolar products.

BARRIER FUNCTION AND HYDRATION 165 Finally, these studies also demonstrate the efficacy of an exogenous lipid mixture on skin moisturization, as assessed by capacitance. An optimal ratio of stratum corneum lipids increases water content in both damaged and normal human skin. Y2 contains 50% of polar lipids, which might retain more water and be superior to physiologic lipids or their mixtures in increasing skin capacitance. The absolute differences in skin hydration between human volunteers and mice are likely due to species differences. Ceramides and fatty acids alone, even in mixtures that disturb barrier recovery (8), will increase skin capacitance in damaged skin (15-18,24). Thus, stratum corneum hydration may be influenced by individual lipids, in contrast to the equimolar or optimized ratios required for barrier homeostasis. Further studies are needed to clarify the relationship of lipids for barrier recovery vs moisturization in this model. In any case, our results suggest that stratum corneum lipids might be useful not only, for skin barrier repair, but also as a skin moisturizer. This extract, and presumably other comparable, naturally occurring mix- tures, may be a cheaper, alternative source of physiological lipids for cosmetic products. ACKNOWLEDGMENT This work was supported by NIH grant AR19098. REFERENCES (1) P. M. Elias and K. R. Feingold, Lipids and the epidermal water barrier: Metabolism, regulation, and pathophysiology, Semin. Dermatol. 11, 176-182 (1992). (2) K. R. Feingold, The regulation and role of epidermal lipid synthesis, Adv. Lipid Res., 24, 57-79 (1991). (3) P.M. Elias, W. M. Holieran, G. K. Menon, R. Ghadially, M. L. Williams, and K. R. Feingold, Normal mechanism and pathophysiology of epidermal permeability barrier homeostasis, ½urr. Opin. Dermatol., 1, 231-237 (1993). (4) K. R. Feingold, M. Mao-Qiang, G. K. Menon, S. S. Cho, B. E. Brown, and P. M. Elias, Choles- terol synthesis is required for cutaneous barrier function in mice, J. ½lin. Invest. 86, 1738-1745 (1990). (5) W. M. Holieran, M. Mao-Qiang, W. N. Gao, G. K. Menon, P.M. Elias, and K. R. Feingold, Sphingolipids are required for mammalian barrier function: Inhibition of sphingolipid synthesis delays barrier recovery after acute perturbation, J. ½lin. Invest., 88, 1338-1345. (6) M. Mao-Qiang, P. M. Elias, and K. R. Feingold, Fatty acids are required for epidermal permeability barrier homeostasis, J. Clin. Invest., 92, 791-798 (1993). (7) G. K. Menon, K. R. Feingold, M. Mao-Qiang, M. Schaude, and P.M. Elias, Structural basis for the barrier abnormality following inhibition of HMG CoA reductase in murine epidermis, J. Invest. Dermatol., 98, 209-219 (1992). (8) M. Mao-Qiang, K. R. Feingold, and P. M. Elias, Exogenous lipids influence permeability barrier recovery in acetone-treated murine skin, Arch. Dermatol., 129, 728-738 (1993). (9) M. Mao-Qiang, B. E. Brown, S. Wu-Pong, K. R. Feingold, and P.M. Elias, Exogenous nonphys- iologic vs physiologic lipids: Divergent mechanisms for correction of permeability barrier dysfunction, Arch. Dermatol., 131, 809-816 (1995). (10) L. Yang, M. Mao-Qiang, M. Taljebini, P.M. Elias, and K. R. Feingold, Topical stratum corneum lipids accelerate barrier repair after tape stripping, solvent treatment and some but not all types of detergent treatment, Br. J. Dermatol., 133, 679-685 (1995). (11) M. W. Holieran, Y. Takagi, K. R. Feingold, G. K. Menon, G. Leglet, and P.M. Elias, Processing of epidermal glucosylceramides is required for optimal mammalian permeability barrier function, J. Clin. Invest., 91, 1656-1664 (1993). (12) G. K. Menon, K. R. Feingold, A. H. Moser, B. E. Brown, and P.M. Elias, De novo sterologenesis in the skin. II. Regulation by cutaneous barrier requirements, J. Lipid Res., 26, 418-427 (1985).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)