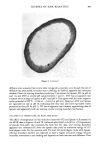
324 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS Experiments to generate dry skin in vitro were done by passing a stream of dry nitrogen over the surface of whole normal pig skin. At timed intervals (0, 6, 24, 48, and 72 hours), the surface was treated with pig sebum, sectioned, stained with Sudan black, and examined microscopically. Slight staining of the stratum corneum could be seen at 6 hours, and by 24 hours the tissue was as darkly stained as in vivo-generated dry skin, with the staining confined to the outer half of the stratum corneum. Identical results were obtained at the 48- and 72-hour time points. Controls which were not exposed to dry nitrogen or which were not treated with sebum remained unstained, i.e., both drying and exposure to sebum were required for the observed staining. Identical results were obtained for human skin. While these tissues did not visually have the appearance of dry skin, cell cohesion is presumably being affected, as it is in vivo, to permit entry of sebum into the stratum corneum. These data and those in Figure 2 indicate that dry skin formation occurs rapidly under harsh environmental conditions. DISCUSSION There are many similarities between pig skin and human skin (21,23-26), including hair density, skin surface structure, epidermal structure, sebum composition, epidermal turnover time, and epidermal lipid composition. Added to the list now is the fact that pigs develop dry skin at low relative humidity this dry skin is visually like that observed on humans. The dry skin responds to treatment with known skin conditioners and to a return to high relative humidity conditions. This animal model is valuable as a tool for rapidly evaluating new skin conditioning actives. Particularly valuable is the ability to test several materials on the same animal, permitting direct comparisons of their efficacies. In contrast to the seasonal availability of human dry skin, pig dry skin can be obtained on a year-round basis with the appropriate controlled environmental facilities. This animal model is also useful as a tool to determine the differences between dry skin and normal skin. The only difference detected by the work done here was an increase in nonpolar lipid in the upper stratum corneum of dry skin. This was determined to be due to migration of surface sebaceous lipid into the cracks and fissures of dry skin, not to any actual change in the epidermal lipid composition. Anderson, Cassidy, Hansen, and Yellin (27) detected an increase in lipid (type undetermined) content of dry skin relative to occlusion-treated dry skin in humans. Also, sebum lipids (28), and triglyceride in particular (29), are elevated in human skin during the winter, the time when dry skin is most prevalent. For pig skin, this lipid difference resulted in a TEWL reduction for dry skin, a reduction which could be eliminated by extraction of the sebaceous lipid. In humans (30), no correlation has been seen between TEWL and skin surface lipid content. Presumably, this lipid is present as a thin film on the skin surface. In pig dry skin, the lipid is deep within the stratum corneum, forming a relatively thick film which could well provide a barrier sufficient to reduce TEWL. We do not believe that the increased sebum lipid content in pig dry skin is a causative factor in dry skin formation. The cracks and fissures of dry skin are simply providing a route for entry of lipid into the stratum corneum where it is retained rather than being lost with surface cells during desquamation. In our histological sections, there were no indications of sebaceous gland density or size differences in dry skin versus normal skin. Since humans routinely expose their skin to surfactant, removing a majority of sebum (28),

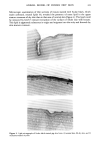
ANIMAL MODEL OF HUMAN DRY SKIN 325 this lipid entry may not occur in human dry skin to the extent that it does in pig dry skin. For dry and visually normal pig skin, no differences in stratum corneum thickness, turnover rate, water-binding capacity, or NMF content could be detected. There were also no differences observed in the viable epidermis. It must be noted, though, that the techniques used will not necessarily reveal subtle differences in the properties examined but certainly indicate the lack of gross differences. The rapid onset of dry skin observed here suggests the lack of involvement of the viable epidermis in the early stages of dry skin formation. However, there may be subtle biological changes in the viable epidermis as a response to the formation of dry skin in the stratum corneum. In humans, there is a variable susceptibility to dry skin formation among individuals under the same environmental conditions. There is certainly an underlying biological/ biochemical explanation for this variability, and a difference in the stratum corneum among individuals certainly exists, resulting in variable resistance to dehydration. The basis for the difference is not known. In pigs, we have not observed variable susceptibility to dry skin formation among individuals. All pigs tested developed dry skin. Anderson et al. (27) observed in humans increases in water-binding capacity, NMF content, and protein for occlusion-treated dry skin relative to untreated dry skin. However, as noted by the authors, the unrealistic total occlusion (from Saran © Wrap) probably increased skin temperature, resulting in an enhanced metabolic rate in the viable epidermis and an increased production of NMF and protein involved in water binding. We are most likely not obtaining total occlusion with petrolatum, and therefore would not encounter the differences reported by Anderson et al. Also, surfactant interactions with skin may be involved in the development of human dry skin. Comparative studies will be necessary to determine the degree of correlation between pig skin and human skin in their normal and dry states. The rapid rate of formation of dry skin on the pig at low relative humidity (Figure 2) suggests that this dry skin is a phenomenon of the stratum corneum and does not require the involvement of viable epidermis or any other insult such as surfactant exposure. Dry skin appears to involve only the upper portion of the stratum corneum (Figure 3). The presence of large surface scales and of sebaceous lipid deep within the stratum corneum indicates defective cell adhesion in dry skin. Since desquamation of surface cells from skin likely involves enzymatic processes (31-32), the aberrant cell adhesion in dry skin may be due to altered enzyme-catalyzed hydrolysis in the stratum corneum. REFERENCES (1) I. H. Blank, Factors which influence the water content of the sratum corneum,J. Invest. Dermatol., 18, 433-440 (1952). (2) P. Flesch and E. C.J. Esoda, Deficient water-binding in pathologic horny layers, J. Invest. Dermatol., 28, 5-13 (1957). (3) K. A. Grice and F. A. Bettley, Skin water loss and accidental hypothermia in psoriasis, ichthyosis, and erythroderma, Brit. Med. J., 4, 195-198 (1967). (4) P. Frost, G. D. Weinstein,J. W. Bothwell, and R. Wildnauer, Ichthyosiform dermatoses. III. Studies of transepidermal water loss, Arch. DermatoL, 98, 230-233 (1968).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)