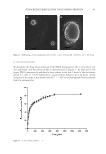
54 JOURNAL OF COSMETIC SCIENCE Since the oxidation rate tapers off at higher fluences, the authors conclude that endogenous flavin chromophores are destroyed by blue light. This flavin resulted in autofluorescence and was subtracted from mouse studies to isolate roGFP measurements. Human mitochondrial versus cytosolic oxidative cell responses mirrored mouse experimental results. Wavelengths in the green light region had no effect on oxidative events in the cytosol or the mitochondria. It was found that blue light cannot produce singlet oxygen as found by analysis of NADH and NADPH fluorescence measurements. Human skin exposed to blue light showed a decrease in autofluorescence measurements. Oxidation and conversion of cellular glutathione to GSSG does not occur very often in healthy cells usually roGFP will remain in a reduced state. This means that if skin cells are under stress conditions, there may be more GSSG existence in the cells. This chain reaction will occur as a result of superoxide and singlet oxygen. The author concludes that blue light was confirmed to contribute to skin aging by a slow and steady exposure to ROS, not by overwhelming cellular antioxidant levels by acute exposure. Melanin and carotenoids both absorb blue light as well as UVA light, but not green light, suggesting that evolutionarily, they are needed to protect the skin from blue light damage as well as UVA. Human skin exposed to normal sunlight levels of blue light showed a disintegration of endogenous flavoproteins (3). As discussed in earlier sections, this can lead to ROS which can signal activation of the MAPK/ERK pathway. Although the glutathione redox state was activated, no long-term blue light oxidative stress is indicated by this specific study as this is a short-term event. Per photon efficacy to activate Table II Blue Light-Induced Events in Comparison with Known UVA and UVB Events Blue light UVA UVB Penetration level Epidermis, dermis Epidermis, dermis Epidermis Main chromophores Flavins, heme, more data needed Flavins, heme, porphyrins, cytochromes Aromatic amino acids, nucleic acids Inflammatory cytokines & inflammatory cytokine-induced MMP IL-1a, MMP1, MMP9, reduction in TNF-alpha, no effect on IL-8 IL-1a, IL-6, IL-8, TNF-alpha TNF-alpha, MMP1, MMP9, MMP3 ROS species known No O2, NO, H2O2, more data needed O2, OH, NO, H2O2 O2, OH, NO, H2O2 DNA impact 8-oxoguanine, T4 endonuclease V (hamster cells in vitro only), more data needed 8-hydroxyguanine and formamidopyrimidines, T4 endonuclease V Pyrimidine dimers Keratinocytes Decrease in viability due to differentiation Decrease in viability Decrease in viability Fibroblasts Reduction in viability for wavelengths below 453 nm Reduction in viability Reduction in viability Wrinkles May lead to wrinkles by loss in dermal integrity Known to lead to wrinkles Known to lead to wrinkles Melanogenesis Through OPN3, may lead to age spots via combination of loss of dermal integrity and melanogenesis in skin types III and above Through p53, known to lead to age spots Through p53, known to lead to age spots

55 POTENTIAL BLUE LIGHT EFFECTS ON AGING the glutathione redox state is about 25% of that of UVA light. It would have been beneficial to repeat this study using times of exposure that were more long term (3). This study indicates that there is short-term oxidative events that occur as a result of blue light exposure. COMPARISON OF BLUE LIGHT SKIN INTERACTIONS WITH UVA AND UVB LIGHT SKIN INTERACTIONS Table II summarizes the differences between the interactions of skin with the different levels of light, as well as the tissue-level effects of the light-activated mechanisms that have been examined. Table III Summary of Previously Examined Data of the Mechanisms Activated by Blue Light Wavelength Fluence Result 410, 412, 415, 419, 420, 426, 453, 460, 470 nm 11 J/cm2 [Nakashima] Reduction in endogenous flavoproteins in vivo 410, 412, 415, 419, 420, 426, 453, 460, 470 nm 50 J/cm2 [Liebel] Hydrogen peroxide generated from human foreheads in vivo 410, 412, 415, 419, 420, 426, 453, 460, 470 nm 65 J/cm2 [Liebel] Increased ROS, no increase in IL-1a, increase in MMP1 and MMP9, Activation of MAPK pathway when in the presence of TNFa 410, 412, 415, 419, 420, 426, 453, 460, 470 nm 130 J/cm2-180 J/cm2 [Liebel] Increased ROS, increase in IL-1a, increase in MMP1 and MMP9, Activation of MAPK pathway when in the presence of TNFa 410 nm 60 J/cm2 [Oplander] Significant reduction in fibroblast viability 412 nm 33 J/cm2 [Liebmann] No effect on IL-8, reduction in keratinocytes and endothelial cells due to differentiation 415 nm 50 J/cm2 [Regazzetti] OPN3 dependent calcium flux causes upregulation of melanogenesis in FSTs III and above 419 nm 33 J/cm2 [Liebmann] Reduction in endothelial cells and keratinocytes due to differentiation 420 nm 60 J/cm2 [Oplander] Significant reduction in fibroblast viability 420 nm 20 J/cm2 [Kleinpenning] No impact on MMP, No impact on inflammatory cytokines, No increase on p53 levels, which increase melanogenesis 426 nm 66 J/cm2 [Liebmann] No effect on IL-8, no reduction in keratinocytes, reduction in endothelial viability 453 nm 18 J/cm2 [Falcone] Decreased IL-1a levels in epidermis following perturbation, increase in TEWL following perturbation, short-lived increase in blood flow following irradiation and perturbation, increase in melanin 72 h after irradiation and perturbation 453 nm 30 J/cm2 [Oplander] NO release in vivo 453 nm 90 J/cm2 [Oplander] No reduction in fibroblast viability 453 nm 100 J/cm2 [Liebmann] No effect on IL-8 460 nm 44 J/cm2 [Nakashima] Increases mitochondrial ROS in keratinocytes 470 nm 3 J/cm2 [Masson-Meyers] No reduction in fibroblast viability 470 nm 55 J/cm2 [Masson-Meyers] No reduction in fibroblast viability 470 nm 110 J/cm2 [Masson-Meyers] Reduction in fibroblast viability 470 nm 220 J/cm2 [Masson-Meyers] Reduction in fibroblast viability
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)