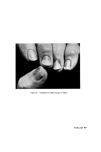
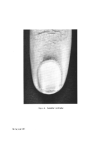
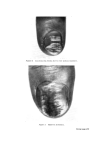
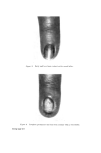
STRUCTURAL ASPECTS OF KERATIN FIBRES 435 notable difference between human hair and wool, apart from the diameter of the fibre, is that human hair (Caucasian) is bounded by a layer of usually six to eight cuticle cells (Figs. 4 and 5), whereas in wool the cuticle is only one to two cells thick. The greater resistance of human hair to chemical attack by many reagents is at least partly due to the thickness of the cuticu- lar layer. Caucasian hair consists of a core of cortical cells mainly of the 'para-type', sometimes surrounded by one to two layers of 'ortho-type' cells (49). However, human hair does not show the same degree of bilateral segmentation as wool. Like wool, human hair is occasionally medullated. Infra-red spectroscopic studies of human hair suggest that the cuticle is made up of the (z and • structures of polypeptide material, while the cortex is composed of both these configurations plus the random coil or amorphous form (50). It is interesting that Negro hair contains a higher proportion of 'ortho- type' cortical cells than Caucasian hair. Also, whereas the cuticle of Caucasian hair is usually six to eight layers thick and covers the whole of each fibre, the cuticle of Negro hair is of variable thickness with six to eight layers at the ends of the major axis of the fibres and diminishing to one to two layers at the ends of the minor axis. Thus, the Caucasian hair approxi- mates to a cylinder, the Negro hair to a twisted oval rod. Negro hair is typically heavily pigmented. By means of the electron microscope, Swift (51) has measured the size of isolated melanin granules and found those from Negro hair to be larger than those from Caucasian and Chinese hair. However, no significant differences in the chemical behaviour and physical properties can be observed between Negro and Caucasian hair (12, 52). Human hair samples give detectable electron spin resonance (ESR) signals at a g value (spectroscopic splitting factor) of 2.003 owing to the free radical property of the melanin granules. The free radical content varies from 4.7 x 10 •6 free spin g-• hair for black hair to 4.6 x 10 x5 for medium brown hair and to 4.3 x 10 •4 for blonde hair (53). During de- pigmentation of black hair with hydrogen peroxide, the intensity of the ESR signal declines exponentially in relation to the treatment time (54). The highest concentration of cystinc in human hair has been found to occur in the 'a' layer and in the exocuficle by an electron histochemical technique (55). It appears that the highest concentration of tyrosine is also in these regions (49). A cell membrane material has been isolated from the ethanol extract of human hair and identified as a lipoprotein (56, 57). Ethanol extraction com- pletely removes this material, since ethanol extracted fibres no longer undergo

436 JOURNAL OF THE SOCIETY OF COSMETIC CI-IEMISTS the characteristic Allw6rden reaction. Furthermore, it has been found that removal of this material by ethanol extraction increases the rate of papain/ bisulphite disintegration of the fibre (58). Examination of sections of metal-coated fibres demonstrated that the cell membrane material is a layer of electron-transparent, non-stainable material approximately 2.5 nm thick on the surface of the fibre. This layer, which is probably the lipid portion of a unit cell membrane, is completely removable by ethanol extraction and may be regarded as originating from the components of the 'epicuticle' described by previous workers. In fact, it has recently been shown (60) that the rate of sorption of n-propanol by wool is increased greatly by preliminary extraction with ethanol, which removes lipid and some protein from the cell membrane complex. This has led to the suggestion that the bimolecular lipid layer of the cell membrane complex, which surrounds each cell within the fibre, presents the major barrier to the diffusion of molecules into the intracellular keratin. END GROUPS OF KERATIN FIBRES In view of the complexity of the histology of wool and hair it is not surprising that the fibres contain a number of both N- and C-terminal amino acids. Methods for quantitative determination of the former are now well defined although techniques for determining the latter are still not as precise. As such groups, as well as the side-chain amino and carboxyl groups, can be involved in reactions with many reagents, they are of considerable importance. Sanger's procedure (61) has been used to identify the N-terminal amino adds. After treatment with 1-fluoro-2,4-dinitrobenzene (FDNB) the fibres are hydrolysed, and chromatographic separation of the substituted amino acids from the hydrolysate shows that, with fibres as dissimilar as human hair and Lincoln, New Zealand Romney, and Australian Merino wools, the terminal groups are always provided by the same seven amino adds, namely, glycine, threonine, valine, alanine, serine, glutamic add and aspartic acid (Table III). Kerr and Godin (65) showed similar end groups to be present in human and horse hair, and H•/hnel (66) found the same end groups in hair, callus, nails, and psoriasis scales. Woodin (67) investigated feather keratin and found the same seven amino acids to be N-terminal and in amounts roughly similar to those of wool.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)