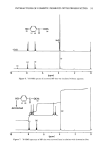
FTIR OF SWEAT GLANDS & ALUMINUM SALTS 121 i I I I I I 4000 3500 3000 2500 2000 1500 1000 500 Wavenumbers (CM '1 ) Figure 7. Subtractions of forearm stratum corneum spectra. ACH-treated minus control (68 hours prehy- drated). ACH treatments were all prehydrated, soaked in 20% ACH for 2 hours, and rinsed for 3 hours in water. Prehydration times for treated samples: A, 66 hours. B, 90 hours. C, 115 hours. that appear as a result of subtraction are different from those in either parent spectrum. Repeat subtractions with different samples must consistently produce similar results. Finally, control subtractions must be performed to determine if artifacts are produced (16). In our forearm stratum corneum subtractions, all four of the above criteria were met. A number of control subtractions, using untreated forearm stratum corneum, were performed, which produced virtually flat baselines at the same scale expansion as in Figure 7. This was due to the sample-to-sample uniformity and good pathlength match described above. Further, the bands which repeatedly appeared in Figure 7 (especially below 1800 cm-•) were at new frequencies not found in untreated stratum corneum. The band near 16 t0 cm-• seems to be due to a lowering of amide I peptide frequency due to coordination between aluminum and carbonyl groups. The literature is replete with examples of metal-amide or metal-amino acid complexation that result in the formation of new bands, mostly between 1600 and 1620 cm -• (l t-13, 18-24). With amino acid complexes, the band is attributed to a metal-carboxylate coordination (usually antisymmetric COO- stretch). The reappearance of the band near 772 cm- • in the subtractions in Figure 7 is particu-

122 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS larly interesting. This band also appeared in the ACH-albumin (Figure 4C) and ACH- sweat protein (Figure 4D) precipitates, which further indicates a strong interaction between ACH and stratum corneum (Figure 7). Although somewhat controversial (18), this band is usually assigned to a metal-coordinated NH 2 deformation mode (11, 12) in metal-amino acid complexes with metal-nitrogen covalent bonds at frequencies between 765 and 790 cm-•. Further, the negative bands near 3300 cm- • in Figure 7 can be assigned to N-H stretch vibrations, demonstrating purturbation around that functional group. These two assignments, plus the shift in the amide B band (also N-H related) described earlier for the plug #2 spectrum (Figure 5A), are good evidence for alu- minum-nitrogen coordination. Whether this coordination occurs directly or via inter- mediate hydroxyl ligands is not distinguished. CONCLUSION Vibrational spectroscopy has been useful in characterizing areas of poral occlusion (plugs) from ACH-treated forearms of two different subjects. Even though the two plug spectra were different, good evidence was found in each for the formation of a complex between aluminum and stratum corneum. One plug spectrum was consistent with ACH-stratum corneum protein interactions plus some free ACH precipitate. The spec- trum from the second plug was a close match with another obtained in vitro by soaking prehydrated axillary stratum corneum in ACH. The protein portion of both plugs was spectroscopically similar to ACH-treated axillary stratum corneum even though the plugs were dissected from forearm biopsies. Perhaps the composition of the keratinized portion within the distal part of any duct is always axilla-like. This ACH-keratin com- plex was distinguished from sweat proteins or ACH-sweat protein precipitates and in- dicates that sweat proteins are not a component of the emphractic material. Binding to keratin was strong and irreversible. It is well established that the upper portion of the intra-epidermal sweat duct is keratin-lined (3,6,26). Epidermal cells migrate laterally toward the ductal lumen and ring the lumen with keratinocytes which actively keratinize and desquamate into the lumen. Thus, within the lumen a supply of keratinized cells is readily available for ACH binding. The hydration state of these cells should be high, a condition which favors aluminum binding. Our spectral evidence, therefore, agrees in effect with the keratin supercontraction hypothesis proposed by Shelley and Hurley (3). A synergistic effect, partially due to the formation of a metal- hydroxide gel emphraxis in the sweat duct [proposed by Relier and Luedders (4)], cannot be totally ruled out. Evidence was found in plug # 1 for the presence of free ACH (band near 1080 cm-1). The spectrum from plug #2, however, showed alu- minum only in the bound state (to keratin), as evidenced by a strong band at 1049 --1 cm It is logical to expect that chemical binding to keratinized ductal walls is required to stabilize a plug and offer resistance to displacement by hydraulic pressure within the duct. This is especially true for ACH, the site of action of which is rather superficially located in the distal portion of the duct within the horny layer. Frictional forces will act to maintain a free forming precipitate (or sweat-protein complex), and such blockage might be expected to float free or be flushed away during firing of a thermally stressed duct.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)