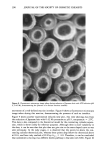
290 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS Figure 8. Fluorescence microscopy image taken during reduction of Japanese hair with ATG solution (pH 9, 0.42 M), demonstrating the presence of a distinct reaction interface. movement of a well-defined reaction interface. Figure 8 shows a fluorescence microscopy image taken during this reaction, demonstrating the presence of such an interface. Figure 9 shows another experimental reduced-time plot, this time showing data from the reduction of Japanese hair with 0.42 M cysteamine at pH 9, temperature = 25øC. This data is also compared to the theoretical model for the contracting cylinder expres- sion, which is shown solely for reference purposes. Although there is more variability in the data, it can be seen that this plot does not give rise to the same shaped curve as was seen previously. In the early stages, it is observed that the points lie above the con- tracting cylinder theoretical plot, whereas these points drop below the theoretical above ot•0.6, and have only reached a•0.8 by t/to. 5 = 4.0. Therefore, it can be concluded that cysteamine is reacting via a different kinetic pathway to that seen with ATG. Figure 10

KINETICS OF HAIR REDUCTION 291 0.9 ß ''' ' '' '• 0.8 o 0.7 ß •v• ^_" o o • 0.6 3 0.5 o sS 0.4 • 0.5 0.2 0.1 0.0 0.5 1.0 1.5 2.0 2.5 5.0 5.5 4.0 t/to.s Figure 9. Reduced-time plot for a series of SFTK experiments using Japanese hair in conjunction with cyste•ine at pH 9, 0.42 M, 25øC. Data are compared to the theoretical cu•e for the contracting •ea model. shows a fluorescence microscopy image of hair reduced by cysteamine, and demonstrates that there is now a more uniform reaction throughout the hair, with no well-defined reaction interface. It may then be concluded that reduction by cysteamine, under these conditions, is giving rise to a reaction-controlled process. Figure 11 shows the untreated ot-vs-time data for reduction of Japanese hair with cysteamine, ATG, and glycerol monothioglycolate (GMT), all at pH 9, 0.42 M. The data demonstrates that under these conditions GMT reacts considerably more slowly than the other two reducing agents however, despite the difference in rates, after normalization it is observed that cysteamine and GMT react via the same kinetic behavior (Figure 12). The usefulness of the reduced-time plots is clearly illustrated in this example, as it is certainly not evident from the raw data that cysteamine and GMT have common kinetic behaviors under these conditions. It should be pointed out that neither of these experimental behaviors can be completely described by any of the deceleratory theoretical models given in Table I. From their deceleratory nature, we may look for the applicability of the geometric, diffusion, or order-based equations however, with the exception of the contracting cylinder model, none are found to apply. The SFTK technique is ideal for looking at the effect of variables such as pH, reactant concentration, temperature, and hair type. It has previously been noted by Wickett that fibers from different individuals may vary significantly in their reactivity towards re- ducing agents. Our experiments have also shown that it is extremely difficult to obtain reproducible results when using conventionally supplied blended hair. It has also been found that analysis of hair from a single individual can also give widely varying results if the fibers are not collected from the same position of the scalp. For example, using fibers that have been removed from an individual's comb or brush can give vastly
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)