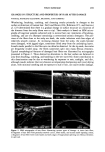
264 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS pendence on fiber diameter (in measurements conducted at 65% R.H.), an increase in the logarithmic decrement for fibers with a larger cuticle content was noted (6). Ap- proximate calculation, based on the assumption that the torsional modulus of the whole, wet hair is 1.8' 10 •ø pascals, yielded values for wet torsional moduli of 2.4 ß 10 •ø pascals and 1.2 ß 109 pascals for the cortex and cuticle, respectively (6). The cortex consists of elongated cortical cells, packed tightly together and oriented parallel to the fiber axis. They contain microfibrils, long uniform filaments which are hexagonally packed into units known as macrofibrils. The microfibrils are composed of highly crystalline material exhibiting a characteristic or-helical X-ray pattern, and are embedded in an amorphous, cysteine-rich matrix. The moisture uptake of hair (14.5% at 65% RH) is confined mainly to the amorphous matrix and results in diametral swelling (16%), with fiber length affected to a minor extent (1.2%) (2). The longitudinal mechanical properties of keratin fibers have been shown to be depen- dent upon temperature, humidity, and time-scale of the experiment and were studied by stress-strain methods (2). These have to be considered in terms of three distinct regions of strain (see Figure 1.). The stress-strain curve up to a few percent strain is referred to as the Hookean region. Further extension occurs with little increase in stress up to about 25-30% and is referred to as the yield region. This constant-stress region is thought to be associated with transformation in the microfibrillar regions. Elongation beyond the yield region leads to a more rapid increase in stress as a function of strain and is called the post-yield region. Many workers attribute the post-yield HOOKEAN YIELD REGION REGION I LOADING CURVE J .......... /////! / /'/•UNLOADING CURVE , o lO STRAIN (%) POST-YIELD REGION I ,, J YIELD POINT AT 20% EXTENS ION ! 20 30 Figure 1. Stress-strain loading and unloading curves characteristic for human hair fibers.

HAIR DAMAGE 265 region to the matrix. The ratios of the moduli of the linear portions of the three regions are approximately 100:1:10. Young's modulus for wool fibers in water was found to vary in the limits 1.7-2.0' 109 pascals, depending upon the rate of strain. As the content of water in the keratin structure is reduced, the stiffness of the fiber increases. In completely dry fibers (0% RH), Young's modulus is augmented by a factor of about 2.7 relative to the same modulus in water. The reference crossectional area in both cases is the value for the wet fibers. It should be noted, however, that the equilibrium stiffness of the fibers (obtained under static conditions and independent of the rate of strain) is independent of the moisture content and corresponds to a value of 1.4 ß 109 pascals (7). It was reported by Feughelman et al. (2) that 6-7 ß 108 pascals of equilibrium Young's modulus value of 1.4 ß 109 pascals are due to coulombic interactions between positively and negatively charged groups in the side-chains of the polypeptides in the keratin structure. Thus, hair fibers tested in aqueous media in the pH range 1-3, the condi- tions under which carboxylic residues are undissociated, showed a 40% decrease in Young's modulus. The mechanical behavior of keratin fibers is often considered in terms of a two-phase model consisting of a water-impenetrable phase of cylindrical rods (microfibrils) ori- ented parallel to the fiber axis, embedded in a water-penetrable matrix phase (8). Ac- cording to the predictions of this model, in longitudinal extension the two phases act in parallel and are equally deformed. In other words, in the wet state, the crystalline phase (microfibrils) contributes considerably to the longitudinal stiffness of the fiber. When the fiber is subjected to torsional stress, on the other hand, and if the matrix is weak- ened by the presence of water, distortion is confined to the matrix. In agreement with the experimental data, the presence of water in the keratin fiber structure should cause a greater reduction in torsional rigidity than in longitudinal stiffness. Based on dynamic mechanical analysis carried out at various humidities, and on conventional mechanical tests at different temperatures in water, estimates have been made of the mechanical contribution of each phase to the equilibrium Young's modulus of 1.4 ß 109 pascals (2). According to these calculations, the contribution of the microfibrils is close to 1.2 ß 109 pascals (about 85% of the equilibrium Young's modulus) and the matrix contribution is of the order of 0.2 ß 109 pascals (only about 15%). PHYSICAL AND PHYSICOCHEMICAL METHODS FOR DETECTION OF FIBER DAMAGE STRESS-STRAIN RELATIONS The measurement of the longitudinal mechanical properties of hair is frequently ap- plied to assess the damaging effects of chemical treatments. Several parameters calcu- lated from stress-strain data were reported to be sensitive to modifications of fibers as a result of cosmetic treatments or environmental degradation. The most commonly used were 20% index, which is the ratio of work required to stretch the fiber by 20% after treatment to the work required to stretch the fiber by 20% before treatment (9), the yield point (stress) at 15% elongation (10), and the tensile strength or extension to break (10). Deem et al. (11) introduced a new parameter, hysteresis ratio (H2o), for the mechanical
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)