PROPERTIES OF KERATIN FIBRES by the work of Bergmann and Sta- ther. Dialanyl cysltine dianhydride, dialanyl cystine and cystine were found to exhibit diminised reactivity towards alkali. Later, Andrews and Andrews demonstrated that cystine phenylhydantoin is much less stable to alkali than is cystine, while Greenstein has reported that cystine cyamidene, crosslinked via salt and cystine link- ages to form grids or sheets. Cohe- sion of the sheets is thought to de- pend on molecular attraction by van der Waals forces. These forces ap- pear to arise in the following way: on quantum mechanical grounds it follows that all molecules possess energy in all states and conse- quently, the nuclei and electrons of NH: C.NH.CH.CH2.S.S.CH2.CH.NH.C: NH I NH,COOH is extremely labile, even in the pre- sence of low concentrations of alkali. Since in proteins the groups at- tached to the cystines linkage are bridged peptide chains, it is reason- able to expect that protein sulphur would be reactive, the reactivity be- ing governed by the strain imposed on the disulphide bond by adjacent chains and linkages. Support for this view has recently appeared in a number of analytical studies of the sulphur and cystine content of wools treated with various, re- agents •ø. The combined cys•dne of wool can be divided into fractions differing in their rate and mode of reaction with sodium bisulphite, al- kalis and formaldehyde. These dif- ferences in behaviour are believed to arise from differences in side chain environments, one fraction being associated with polar side chains, e.g., salt linkagesl, while the other fraction is associated with non-polar side chains. So far, then, the molecular struc- ture of the wool fibre is pictured as a series of long polypeptide chains COOH NH2 atoms undergo some kind of relative vibration. This causes the forma- tion of temporary dipoles which are able to induce in other molecules dipoles in phase with themselves. As a result there is a net attraction between the molecules which is con- sidered to be the major factor con- tributing to the van der Waals forces. Additional linking of adjacent sheets almost certainly occurs by co-ordination of peptide linkages with the formation of hydrogen bonds: \ / \" NH ......... CO NH / -,, CO NH ......... CO 'x / CHR CHR CHR / ,,, / NH ......... CO NH CO NH ......... CO / \ / Further details of the molecular structure of woo] arose from correla- tion of the elastic properties of the fibre with the results of X-ray work TM. Earlier, it had been con- 161

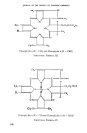
JOURNAL OF THE SOCIETY OF COSMETIC CHEIlilS'rS cluded in the case of cellulose and silk that the chain molecules con- stituting the fibre crystallites are as fully extended as possible •ø. This view was in agreement with the fact that on stretching fibres (1) the X- ray diffraction patterns remain un- changed (2) fibres do not recover their original length completely when the stretching force is re- moved. Stretching of cellulose and silk was, therefore, considered to occur by internal slipping of chain bundles over one another. Wool, on the other hand, does not behave in this way. Wool fibres in water ex- hibit long range elasticity, with the property of recovering their original length when the stretching force is removed. Furthermore, X-radio- grams of normal wool (a-keratin) are different from those of natural silk. However, as wool or hair is stretched, the normal X-ray photo- •aph gradually disappears and is replaced by a new one (/9-keratin) which is similar to that given by natural silk. It was concluded from this that stretched wool is built from fully extended polypeptide chains. When wool fibres which have been stretched in water at ordinary tem- peratures are released in water, they quickly recover their original length, and, at the same time, the normal X-ray pattern returns. This process can be repeated many times, show- ing that the transformations are re- versible. Since stretched wool was considered to consist of fully ex- tended protein chains, it was con- cluded that the unstretched fibre is built of the same chains in a regu- larly folded state. This was the position in the early 1930's, since which time much discussion has centred round the nature of the fold. The original hexagon fold of keratin: -- was later shown to be incapable of providing adequate space for accom- modating the amino acid residues, and the model was accordingly re- vised as show below•a: I co x.H H C--NH--CO--C R Rx NH I co RCH NH I co H Hx x C---CO--NH--C NH R R I It has been suggested by other workers, however, that a- and /•- keratin consist of many different types of fold, and it is believed that 162
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)