30 JOURNAL OF COSMETIC SCIENCE is clearly only fragmentary. More research will be necessary to reveal its true identity and to gain an understanding of the nature of its molecular interactions with the adjacent [B-layers. THE [3-LAYERS Each [B-layer of the CMC is associated with an adjacent cuticle cell surface and, since not one of the many heavy-metal staining procedures for the TEM results in its intensifi- cation, it has always been supposed to consist mainly of saturated lipids (see below). Since the [B-layers seemed to contour each cuticle cell, it was not unreasonable to suppose they also extended to cover the outer cuticular surface of the fiber. Direct visual evidence for such a surface [B-layer was first provided by Swift and Holmes (33) from TEM observations of human head hairs, and this has been confirmed recently by Jones and Rivett (1). It is convenient to discriminate between two types of [B-layer (cf. Figure 2). The upper [B-layer is that attached to the A-layer on the aspect of each cuticle cell sheet facing the outside of the fiber, both within the CMC between the cells but also extending across the free outer surface of the hair. Conversely, the lower [B-layer is on the side of each cell facing the fiber cortex and attached to the inner layer. Each cuticle cell sheet is very thin compared with its in-plane dimensions, and so one expects a relatively sharp cut-off at the edges of each cell between the two types of [B-layer. In some hairs one finds under the TEM that the two types of [B-layer are of approximately the same thickness within the CMC, but in other hairs the upper [B-layer appears to be slightly thicker than the lower [B-layer (11). Such a difference undoubtedly signifies variations in the lipid com- position between the two layers (see below). It is noteworthy in the context of the later discussion that, following the rigors of cutting thin sections of hairs for the TEM, one often finds that splits have developed in the CMC along the line of the upper [B-layer but rarely in the lower [B-layer (34). This highlights the possibility of specific mechanical weakness associated with the upper [B-layer. On the other hand, it remains unresolved by TEM observation whether such failure occurs cohesively within the bulk of the upper [B-layer, or adhesively at one or other of its interfaces with the A-layer or with the central 8-layer. COVALENTLY LINKED FATTY ACIDS IN THE CUTICULAR B-LAYERS At a point in time when scientists thought they knew most of the basic chemistry of mammalian keratin fibers, the remarkable discovery was made of fatty acids covalently attached to the outer surfaces of wool fibers (35) and contained within the bulk of almost all mammalian keratin fibers (36,37), apart from the lowest order of Mammalia, the monotremes (38). What followed was a flurry of research activity, led principally by the Australians, toward understanding the detailed chemistry, modes of attachment, and location of these fatty acids. The excellent up-to-date review on the subject by Jones and Rivett (1) is a fine tribute to the memory of the late Don Rivett and the part he played in this pioneering work. Selected information is now drawn from this review for further discussion.

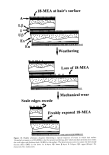
HUMAN HAIR CUTICLE 31 Current evidence points to the covalently linked fatty acids being contained within the [•-layers of the CMCs of both the cuticle and cortex where, in the native fiber, they undoubtedly exist in admixture with loosely bound lipids. What distinguishes the two types of cellular CMC from each other is that those in the cuticle contain the unusual methyl-branched saturated fatty acid, 18-methyl eicosanoic acid (18-MEA) (cf. Figure 4), as a major covalently bound component, whereas only traces of this particular fatty acid are encountered in the cortex (39). 18-MEA comprises in the region of 50% w/w of the bound fatty acids in the cuticle, with palmitic (C16:0) and oleic (C18:1) acids as the next most abundant (39). Some controversy still exists about the precise mode of attachment of these fatty acids, but the balance of chemical evidence favors their being linked via a thioester to the cysteine residues of a supporting protein (40). Protrey and Ferguson (41) established that in skin the anteiso-fatty acids with odd numbers of carbon atoms (as is the case for 18-MEA) are specifically synthesized from the precursor amino acid, isoleucine. A key step in the biosynthetic pathway involves a branched-chain 2-oxo acid dehydrogenase but, through a genetic defect, this enzyme is absent in the rare inherited disorder known as maple syrup urine disease (MSUD) (42). In this disease 18-MEA is not synthesized, as is indicated by the virtual absence of this fatty acid from the patient's hair (43). TEM observations of hairs from MSUD patients show that the cuticle's upper [•-layer was defective, and this has led to the conclusion that in normal hair 18-MEA is located principally in this layer (44). THE NUMBER OF MONOLAYERS OF 18-MEA IN HAIR Valldorf (45) has determined the 18-MEA content of European hair of 25-cm length. As might be expected from the gradual mechanical attrition of cuticle at the hair's surface (4), its 18-MEA content decreases gradually from the root to the tip. Linear regression of his results provides an indication that the 18-MEA content of the root end is 407.9 lag per gram of hair. This corresponds reasonably well with there being just one mono- layer of 18-MEA between each cuticle cell, a monolayer at the hair's outer surface, and the likelihood of a further monolayer at the surface of the cortex (cf. Appendix 1). One presumes that most of this is contained within the cuticle's upper [•-layer and that there is none in the lower [•-layer. PHYSICAL PROPERTIES OF 18-MEA The terminology "cell membrane complex" creates the false impression of this structure being analogous to the cell membranes of living cells, a misinterpretation arising out of their similarity of structure in the TEM, coupled with their common location at the ' ' 2.39 nm ,,,CH2 CH2 CH2 CH2 CH2 CH2 _CH2 CH2 .CH2 ....COOH H3C CH CH 2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 I CHa 18-methyl eicosanoic acid Figure 4. The structure of 18-methyl eicosanoic acid. The approximate length of the molecule from the anreiso-terminus ro the carboxyl carbon is shown and was calculated by Jones and Rivert (1).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)