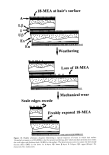
34 JOURNAL OF COSMETIC SCIENCE been identified among the other fatty acids during chemical analysis, but it remains an attractive possibility that this type of degradation might account for the decrease in hydrophobicity of the hair's surface in going from root to tip (45). CONFORMATION OF LIPIDS IN THE [•-LAYERS It would be useful to know accurately the thickness of the cuticular lB-layers as a starting point for considering the conformational disposition of the lipids within them. From TEM observations of transversely sectioned hairs, various authors have quoted the thick- ness of the lB-layers of the CMC as being somewhere between 2.5 and 4.0 nm. This is unfortunate because the lower value would be consistent with a monolayer of an ex- tended lipid chain such as 18-MEA, whereas the higher value is what might be expected for a typical lipid bilayer. One should not underestimate the difficulty of using the TEM to measure the thickness. Even assuming accurate calibration of the microscope at the high image magnifications required for the job, the main problem is that the lB-layer is contained within a section of the hair whose thickness is approximately 20 times that of the lB-layer. Even small deviations in planarity of the lB-layers within the section, or small angles of tilt in the plane of the lB-layer from being parallel to the electron beam, will serve to confound the relevance of the measurements. The problem might be resolved by using a TEM equipped with a stage goniometer to tilt the sections incre- mentally about the plane of the lB-layer, but such concerted studies have not been reported. As indicated above, great uncertainty exists as to whether the lipids in individual lB-layers of the CMC are monolayers or bilayers. Current evidence points to a monomo- lecular layer of 18-MEA being attached to the A-layer of each cuticle that constitutes the whole or part of the upper lB-layer. One presumes that the other covalently linked fatty acids, which occur in cuticle in amounts approximately equal to that of the 18-MEA, are attached to the inner layer and constitute the whole or part of the lower lB-layer. Stoichiometrically this accounts for all the covalently linked fatty acid normally present in the cuticle, and so one must exclude the possibility of there being further monolayers of covalently linked fatty acid attached to the surfaces of the 8-layer that might other- wise form bilayers with the existing covalently attached lipids in the upper and lower lB-layers. What remains unresolved are the types, amounts, and disposition of free lipids within the cuticular CMC. Some of these might be located within the body of the 8-layer, but then the ease with which this component can be stained with heavy metal compounds would seem to belie a greater level of hydrophilicity than would be the case for a structure containing significant amounts of lipid. More likely it is that these other lipids form pseudoclassic bilayers with the covalently bound lipids of the upper and lower lB-layers, the polar ends of these free lipids interfacing with the surfaces of the 8-layer. More research will be required to test the veracity of such a model. Uncertainty also exists about the thickness the upper lB-layer presents at the outer surface of degreased fibers. From the TEM of transverse fiber sections, thicknesses of from 2.5 nm (33) to 6.0 nm (1) have been quoted. To add yet further to this uncertainty, Ward et •/. (8) determined by X-ray photoelectron spectroscopy that the thickness of the hydrocarbon layer afforded by 18-MEA at the surface of wool fibers is only a matter of 0.9 nm. Zahn et •l. (18) presented a model in keeping with this latter thickness, in

HUMAN HAIR CUTICLE 35 which surface chains of 18-MEA were folded back on themselves. This folded-chain model might be conformationally adopted as a mean for burying the intense hydropho- bic anteiso terminus of 18-MEA when solvent-extracted fibers are exposed to water. On the other hand, one must not lose sight of the fact that animal hairs are normally covered with sebaceous excretions where the 18-MEA chain can be expected to be expected to intercalate with those lipid chains. Indeed, the highly mobile anteiso terminus of 18- MEA could under such circumstances facilitate the spreading of these extraneous lipids across the fiber surface by reducing interfacial shear strength. Considerable controversy thus currently exists as to the actual thickness of the surface [3-layers and the attendant conformation of 18-MEA. This author believes these uncertainties might be resolved by concerted atomic force microscope (AFM) investigations of the surfaces of solvent- extracted hairs. He envisages that the thickness of the external 18-MEA layer might be determined from force-distance plots for the vertical penetration of the surface lipid by a sharply profiled AFM probe tip, where the lower end of the lipid chains would be sensed by a rapid rise in resistance as the crosslinked proteinaceous supporting layer is reached. He also anticipates that by AFM it will be possible to gain considerable insight into the molecular rheology of the surface 18-MEA by making use of a transversely oscillating probe tip. THE PROTEIN SUPPORTING 18-MEA Current evidence is that 18-MEA at the cuticle surface is attached via thioester linkages to the cysteine residues of an underlying layer of protein. The connecting cysteine residues would need to be presented to the outer surface of this protein in sufficient density to maintain rnonornolecular coverage of the 18-MEA. One therefore expects these cysteines to be in pseudohexagonal array and presented at an average center-to- center separation of the order of 0.936 nm to satisfy the monolayer condition (N.B.: 0.936 nm is the diameter consistent with the end-on area for each 18-MEA molecule-- cf. Appendix 1). Negri et •l. (51) have already suggested that the protein could perhaps be in the form of a [3-pleated sheet in which extended protein chains ([3-strands) lie parallel and hydrogen-bonded to each other. Such proteins offer the opportunity for amino-acid side chains to be presented at one side of the sheet at intervals of between 0.64 and 0.68 nm along each strand, for strands 0.48 nm apart. The existence of amino- acid side chains on the other side of the sheets at the same density means that the protein wouldn't need to possess a particularly high cysteine content to satisfy bonding with 18-MEA. On the other hand, a high cysteine content would be expected to satisfy the needs for intermolecular disulphide bonding on the obverse side of the sheet (i.e., between adjacent [3-strands within the sheet and with other underlying cysteine-rich proteins). Attempts to isolate and analyze fragments of the proteins still attached to 18-MEA have so far proved unsuccessful (45,50). This is unfortunate because such an approach would provide the potential for identifying the corresponding gene and for obtaining a complete amino acid sequence for the protein to which 18-MEA is attached. DIFFERENTIAL FRICTION AND ITS ORIGIN A property of human head hairs, as for all undamaged mammalian hair fibers, is that the coefficient of friction for rubbing their surfaces in a root-wise direction is significantly
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)