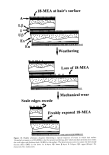
44 JOURNAL OF COSMETIC SCIENCE 5. Differential friction that makes the hair on the head naturally disentangling and aligned and aids in the ejection of scalp surface detritus. 6. An upper [3-layer that, by its liquid-like behavior and relative mechanical weakness, ensures that fresh 18-MEA is presented to the hair's outer surface as mechanical attrition of the cuticle takes place. This aids in the preservation of differential friction and the associated advantages. APPENDIX 1 NUMBER OF LAYERS OF 18-MEA IN THE CUTICLE Here we compare Valldorfs (45) analyses for the 18-MEA content of root-end hair with theoretical estimates and derive an estimate of the number of layers of 18-MEA in the cuticle. Our initial assumptions are that the European hairs Valldorf used correspond to circular rods of 70 t•m average diameter (66) and that the root-end of each hair has ten cuticle cell layers at its periphery. If we consider initially that the entire outer surface of each hair is covered with a monolayer of 18-MEA, then the mass of 18-MEA in a given length of the fiber will be given by (surface area of hair) x (moleclar weight for 18-MEA) (surface area of each 18-MEA molecule) x (Avagadro's number) The surface area of the hair can be readily calculated if we know its diameter (70 pm) and length (for initial convenience we will choose 10 mm). The molecular weight of 18-MEA is 326, the surface area occupied by the anteiso-terminus of each 18-MEA molecule is 68.89 x 10 -2ø m 2 (45), and Avagadro's number (i.e., the number of molecules in one gram mole) is 6.025 x 1023. Upon substituting these quantities in the equation, the theoretical amount of 18-MEA covering the outer surface of a 10-mm length of the hair will be 1.728 x 10 -9 g. The mass density of hair is normally accepted to be 1.30 g cm -3, and on this basis, the mass ofa 10-mm length of 70-pm hair is almost exactly 50 pg. A monolayer of 18-MEA therefore corresponds to 34.6 pg of 18-MEA per gram of hair. Linear regression of Valldorf's (45) practical analyses for the amount of 18-MEA with distance along a hair sample yields a figure of 407.9 pg of 18-MEA per gram of the hair's root end. This corresponds to 11.8 theoretical monolayers of 18-MEA in the hair cuticle. Given our normal expectation of ten layers of cuticle at the root-end of human hair, this is in reasonable accord with a total of eleven monolayers of 18-MEA comprising one layer on the outer-facing surface of each cuticle cell, one monolayer in each of the nine intercuticular cell membrane complexes, and a further layer at the surface of the cortex. Interestingly, the results of Negri et al, (51) for the 18-MEA content of wool (2.3 theoretical layers) are consistent with this general model. Negri et al.'s original results were interpreted on the basis of a monolayer of 18-MEA around the entire perimeter of each cuticle cell. At any given point along a wool fiber there are either one or two layers of cuticle as provided by a single layer of scales overlapping for an average of approximately 0.3 of their length. The 2.3 theoretical layers can thus be alternatively ascribed to one monolayer at the surface of the fiber, one monolayer at the surface of the

HUMAN HAIR CUTICLE 45 cortex, and 0.3 of a monolayer layer associated with the intercuticular cell membrane complex. REFERENCES (1) L. N. Jones and D. E. Rivett, Role of 18-methyleicosanoic acid in the structure and formation of mammalian hair fibres, Micron, 28, 469-485 (1997). (2) J. A. Swift, Human and other animal hairs: Morphological and functional comparison, Retinoids, 13, 114-119 (1997). (3) M. L. Garcia, J. A. Epps, and R. S. Yare, Normal cuticle wear patterns in human hair, J. Soc. Cosmet. Chem, 29, 155-175 (1978). (4) J. A. Swift and A. C. Brown, The critical determination of fine changes in the surface architecture of human hair due to cosmetic treatment, J. Soc. Cosmet. Chem., 23, 695-702 (1972). (5) J. A. Swift, "Morphology and Histochemistry of Human Hair," in Formation and Structure of Human Hair, P. Jollbs, H. Zahn, and H. H6cker, Eds. (Birkh•/user Verlag, Basel, 1997), pp. 149-175. (6) J. A. Swift and B. Bews, The chemistry of human hair cuticle. Part 3. The isolation and amino acid analysis of various sub-fractions of the cuticle obtained by pronase and trypsin digestion,.]. Soc. Cosmet. Chem., 27, 289-300 (1976). (7) J. A. Swift, Minimum depth electron probe X-ray microanalysis as a means for determining the sulphur content of the human hair surface, Scanning, 2, 83-88 (1979). (8) R.J. Ward, R. E. H. Willis, G. A. George, G. B. Guise, R. J. Denning, D. J. Evans, and R. D. Short, Surface analysis by X-ray photoelectron spectroscopy and static secondary ion mass spectrometry, Textile Res..], 63, 362-368 (1993). (9) J. Shao, D. C. Jones, R. Mitchell, J. C. Vickermann, and C. M. Cart, Time-of-flight secondary-ion mass specrromerric (Tof-SIMS) and X-ray photoelectron spectroscopic (XPS) analyses of the surface lipids of wool,.]. Textile Inst., 88, 317-324 (1997). (10) B.C. Powell and G.E. Rogers, "The Role of Keratin Proteins and Their Genes in the Growth, Structure and Properties of Hair," in Formation and Structure of Human Hair, P. Jollbs, H. Zahn, and H. H6cker, Eds. (Birkh•/user Verlag, Basel, 1997), pp. 59-148. (11) G. E. Rogers, Electron microscope studies of hair and wool, Ann. N. •5 Acad. Sd., 83,378-399 (1959). (12) J. M. Gillespie, "The Structural Proteins of Hair: Isolation, Characrerisarion and Regulation of Bio- synthesis," in Physiology, Biochemistry and Molecular Biology of the Skin, L. A. Goldsmith, Ed. (Oxford University Press, Oxford, 1991), Vol. 1, pp. 625-659. (13) B. J. Jenkins and B. C. Powell, Differential expression of genes encoding a cysteine-rich keratin family in the hair cuticle,.]. Invest. Dermatol., 103, 310-317 (1994). (14) P.J. Mackinnon, B.C. Powell, and G. E. Rogers, Structure and expression of genes for a class of cysreine-rich proteins of the cuticle layers of differentiating wool and hair follicles,.]. Cell Biol., 111, 2587-2600 (1990). (15) J. A. Swift and B. Bews, The chemistry of human hair cuticle. Part 2. The isolation and amino acid composition of the cell membranes and A-layer,.]. Soc. Cosmet. Chem., 25, 355-366 (1974). (16) K. Yoneda, M. Akiyama, K. Morita, H. Shimizu, S. Imamura, and S.-Y. Kim, Expression of trans- glutaminase 1 in human hair follicles, sebaceous glands and sweat glands, Br..]. Dermatol,, 138, 37-44 (1998). (17) M. Nienhaus, Beltrag zur Enzymatischen Hydrolyse yon Keratinfasern und lhren Komponenten, Ph.D. thesis, Rheinisch-Westoealischen Technischen Hochschule, Aachen, Germany (1981). (18) H. Zahn, H. Messinger, and H. H6cker, Covalenrly linked fatty acids at the surface of wool: Part of the "cuticle cell envelope," Textile Res..]., 64, 554-555 (1994). (19) I. K. Allwt3rden, Die eigenschaften der schafwolle und eine neue untersuchungsmethode zum nach- weiss gesch•/diger wolle auf chemischen wege, Z. Angew. Chem., 29, 77-78 (1916). (20) J. H. Bradbury and J. D. Leeder, Keratin fibres. Part 6. Mechanism of the Allw6rden reaction, Aust. .]. Bid. Sci., 25, 133-138 (1972). (21) P.M. Steinerr and L.N. Marekov, The proteins elafin, filaggrin, 1oricrin, and small proline-rich proteins 1 and 2 are isopeptide cross-linked components of the human epidermal cornified cell envelope,.]. Biol. Chem., 270, 17702-17711 (1995). (22) R. H. Rice, V.J. Wong, and K. E. Pinkerton, Ultrastructural visualisarion of cross-linked protein features in epidermal appendages,.]. Cell Sd., 107, 1985-1992 (1994).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)