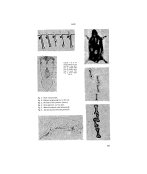
HAIR Figures I to 6 are shown herewith: top left, Fig. I top right, F•g. 2 middle left• Fig. 4 middle right, Fig. 5 lower left, Fig. 3 lower right, Fig. 6. Fig. I. Hair replacement, Fig. 2. Pattern of growing hair in the rat. Fig. 3. Primitive hair pattern (otter), Fig. 4. Hair patterns on lion skin. Fig. 5. Medulla stained with Safranine B. Fig. 6. Cortex stained with Kiton Red G. 161

JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS to the pattern and relative thickness of the medulla. (3) Examination of cuticular scale pattern. Knowledge 'of fibre characteristics resulting from such examination is of value to biologists, taxidermists, archaeologists, forensic scientists, while the microscopic examination of human hair is of importance in certain pathological scalp conditions. Histochemical studies carried out on fibre sections have yielded valuable information about the physico-chem- ical properties of cuticle, cortex and medulla 9'•ø (Figures 5 and 6), the significance of which in the formula- tion of permanent waving solutions has been described elsewhere. TM HAIR PIGMENTATION The pigmentation of hair has been studied extensively by geneticists, typical investigations being those of Onslow, Wright and Koller." Such work clearly shows that pigment formation results from the action of enzymes on a basic chromogen to give the basic colours, viz.: black, and red or yellow, the intensity and dis. tribution of which are governed by factors chiefly genetic in origin. The resulting coat patterns in animals behave as genetic characters while subject to variation which can often be modified by selection. In albin- ism (recessive whiteness) there is a complete failure to inherit the enzyme system, while in dominant whiteness an enzyme inhibitor is present and prevents formation of melanin. These views explain the changes in coat colour which can be produced experimentally in mature animals by shaving, or by local subjection to low temperatures, since the formation of enzyme and chromo- gen appears to be controlled by skin temperature, which constitutes an intermediate link in the chain of genetic causation. The absence of pigment from the coats of snow- field fauna doubtless results from a similar suppression of the chemical reactions leading to melanin forma- tion. The chemistry of melanin forma- tion has been examined by Raper, et aI., •3 who have described three stages in the oxidation of tyrosine to the black pigment. In the first stage tyrosine gives rise to a red pigment, which is followed in the second stage by formation of a colour- less substance. In the third stage the colourless substance is oxidised to melanin. The first stage is the only one requiring the presence of the enzyme (tyrosinase), and, since the second stage occurs in an atmosphere of nitrogen, it cannot be oxidative. At the end of the second stage the presence of 3:4 dihydroxyphenyla- lanine, 5: 6 dihydroxyindole and 5: 6 dihydroxyindole-2-carboxylic acid can be demonstrated, and in order to explain these facts Raper put forward the scheme shown on the next page. The mechanism of conversion of 5:6 dihydroxyindole and its 2-car- boxylic acid into melanin is obscure, and the structure of the pigment remains unknown. This classic work of Raper was followed by study of melanin precursors, first by Burton 162
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)