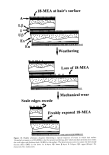
HUMAN HAIR CUTICLE 25 compounds. According to the type of stain used, some knowledge of the chemistry of the hair's microscopic components has been deduced (5). Additionally, TEM examination of sections treated with various proteolytic enzymes, coupled with chemical analyses of physically isolated cuticle treated with these same enzymes, has provided a valuable insight into the amino acid composition of the cuticle's subcomponents (6). Other methods that have aided chemical analyses of components close to the hair's cuticular surface are X-ray microanalysis (7), electron spectroscopy for chemical analysis (ESCA) (also known as X-ray photoelectron spectroscopy, or XPS) (8), and secondary ion mass spectroscopy (SIMS) (8,9). The cloning and DNA sequencing of genes is now providing useful information about the amino acid sequences of proteins present in the cuticle (10). On the other hand, it is but speculation as to which subcomponents of cuticle contain these proteins, and uncertainty remains as to their overall conformations and roles. Each cuticular sheet consists of a series of yet thinner sub-sheets and is separated from its neighbors by an even thinner laminated cell membrane complex (CMC) (cf. Figure 2). A detailed description of each subcomponent now follows. THE A-LAYER First identified in the cuticle of human hair and wool and named by Rogers (11), this proteinaceous component, located on the outer-facing aspect of each cell, is of constant thickness (ca. 110 nm) within each cell. The thickness is more or less the same both within and between different animal species. Its cystinc content, at approximately 1 in every 2.7 amino acid residues as 1/2-cystinc (7), is exceedingly high and as such would seem to fall within the class of so-called ultrahigh sulphur (UHS) proteins commonly encountered in protein analyses of mammalian keratin fibers (10,12). No accurate amino acid analysis is available for an isolated A-layer fraction. On the other hand, the proteins contained therein could well be similar or identical to one or other of the UHS proteins from cuticle, the amino acid sequences for which have been established by the DNA sequencing of the corresponding genes (10,13,14). Upper [3-layer at hair's surface • A-layer•escedgeFractured f :"" '":: ! ' :' :"' ::" :'"'"': :'" :"" , Exocuticle ....
Lower :.... Cell • [3-layer • :5 '• '/' Endocuticlayer membrane. j 6, la • • ,:• ,•..,,• ii ii1•1111 ompex II II1111 (CMC) • Upper '. .... ".:.'. ':':.•: •_•_ayer Figure 2. Highly schematic diagram showing the longitudinal internal structure of the cuticle dose to a surface scale edge, with respect to the immediately underlying cuticle sheet. The scale edge represents one that has been mechanically abraded, thereby exposing the lameliar subcomponents at the hair's surface.

26 JOURNAL OF COSMETIC SCIENCE That the A-layer was relatively resistant to digestion by mixtures of papain and dithio- threitol (15), under conditions in which almost all the other hair's components dissolve, was the first indication that perhaps this structure contained isodipeptide crosslinks in addition to the large amounts of cystine disulphide crosslinks already known to be present. Such crosslinks occur as a protein intermolecular peptide bond, ½-amino-(•/- glutamyl) lysine, and their formation between lysyl and glutaminyl side chains in the follicle is facilitated by the calcium-dependent enzyme, transglutaminase (cf. Figure 3) (16). Fractions of cuticle derived by papain/dithiothreitol digestion, and containing the CMC and significant residues from the A-layer, were shown by Nienhaus (17) to possess isodipeptide at a level of ca. 70 pmol/g. Later Zahn eta/. (18) estimated a concentration of 2.5 mole% of isodipeptide for the enzyme-resistant material, based on the assumption that 50% of the 5 mol% of the protein's lysine residues had participated in isodipeptide formation. What remains uncertain is the distribution of these isodipeptide-rich pro- teins within the cystine-rich A-layer or indeed within the cuticle as a whole. An attractive notion is that they are concentrated in a superficial subcomponent of the i co Glutamine sine co I I HC"-- C-"- C-'- C'--- N H2+ H2N•-C•C•C---C CH I H H0 H H H H2 I NH NH ! ii ! ! ! ! I I I Transglutaminase +Ca 2+ (-NH3) C---- C • C-'----N---- C---- C--- C---- C---- CH isopeptide linkage H2 H2 H2 NH ! s-ami no-(l,-gl utamyl)lysine Figure 3. The formation of isopeptide intermolecular crosslinks. This occurs in some hair proteins by the action of transglutaminase upon spatially proximate glutamine and lysine sidechains.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)