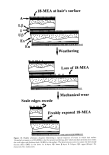
HUMAN HAIR CUTICLE 29 and is between 10 and 40 nm thick. The term "inner layer" seems to have been first coined by Orfanos and Ruska (28). Its appearance under the TEM, its high cystine content, and its susceptibility to protease attack all mimic the exocuticle. The proteins it contains are therefore probably identical to those of the exocuticle. Functionally it provides rigid support for the lipids of the lower [3-layer (see below). THE CELL MEMBRANE COMPLEX (CMC) This thin multicomponent layer separates all cuticle cells from each other. The detailed internal structure and today's accepted nomenclature for the components were first presented by Rogers (11) following his seminal observations with the TEM of stained sections of hair and wool. The CMC is more or less planar and separates cuticle cells from each other. It is of relatively constant thickness (in the range 25 to 28 nm). PTA staining provides excellent contrast in the TEM for the CMC to exhibit a tram-line appearance of two unstained layers ([3-1ayers) of between 2.5 and 4.0 nm thickness, separated by an intensely stained layer (8-layer, 8-band, intercellular cement or "glue") 15-18 nm thick. The CMC is generally considered to embrace the two [3-1ayers and the intervening 8-layer, although in the fullness of time, it might be considered appropriate to include other thin peripheral layers of protein. THE 8-LAYER Jones and Rivett (1) have shown in sections of human hair, pretreated by Rogers' (11) thioglycollic acid/osmium tetroxide procedure and then post-stained with UA/LC, that the 8-layer contains a central lamina of ca. 5 nm thickness and of lesser staining intensity. Great uncertainty remains about the chemical composition of the 8-layer and its substructures on account of the difficulty of isolating it chemically unchanged and uncontaminated by components from other parts of the hair shaft (29). There is ample evidence from electron histochemical observations that the 8-layer does not contain cystine (5), but one needs to keep an open mind about whether it's mainly proteinaceous or whether it might be polysaccharidic or a mixture of the two, like a glycoprotein. This author inclines to the latter viewpoints principally because of the resistance of the cuticular 8-layer to the action of the highly aggressive protease agent papain/dithiothreitol in treatments either of the bulk fibers or thin sections of them (15). The 8-layer stains with the periodic acid/silver methenamine procedure, a well-known method for polysaccharides in which vicinal-diols are oxidized by periodate to aldehydes, which in turn result in the deposition of granular silver with the silver reagent (5). Intense staining of the 8-layer seen with PTA can also be explained as due to the ionic interaction at low pH of the phosphotungstate trivalent anion with the protonated hydroxyls of a polysaccharide (5). There is also electron histochemical evidence for the presence of free amino groups that also bind PTA (30). Yet further, Orwin (31) in TEM investigations of sections of wool follicles observed a polysaccharidic coating on all the cells, and it is not unreasonable to suppose this material becomes incorporated as the principal component of an intercellular "glue" in the 8-layer. Allen eta/. (32) discovered glycoproteins in the residues extracted from a variety of keratin fibers (including human hair) with 98% formic acid, a reagent that is thought to remove material, albeit probably not specifically, from the CMC. Our knowledge of the chemical makeup of the 8-layer

30 JOURNAL OF COSMETIC SCIENCE is clearly only fragmentary. More research will be necessary to reveal its true identity and to gain an understanding of the nature of its molecular interactions with the adjacent [B-layers. THE [3-LAYERS Each [B-layer of the CMC is associated with an adjacent cuticle cell surface and, since not one of the many heavy-metal staining procedures for the TEM results in its intensifi- cation, it has always been supposed to consist mainly of saturated lipids (see below). Since the [B-layers seemed to contour each cuticle cell, it was not unreasonable to suppose they also extended to cover the outer cuticular surface of the fiber. Direct visual evidence for such a surface [B-layer was first provided by Swift and Holmes (33) from TEM observations of human head hairs, and this has been confirmed recently by Jones and Rivett (1). It is convenient to discriminate between two types of [B-layer (cf. Figure 2). The upper [B-layer is that attached to the A-layer on the aspect of each cuticle cell sheet facing the outside of the fiber, both within the CMC between the cells but also extending across the free outer surface of the hair. Conversely, the lower [B-layer is on the side of each cell facing the fiber cortex and attached to the inner layer. Each cuticle cell sheet is very thin compared with its in-plane dimensions, and so one expects a relatively sharp cut-off at the edges of each cell between the two types of [B-layer. In some hairs one finds under the TEM that the two types of [B-layer are of approximately the same thickness within the CMC, but in other hairs the upper [B-layer appears to be slightly thicker than the lower [B-layer (11). Such a difference undoubtedly signifies variations in the lipid com- position between the two layers (see below). It is noteworthy in the context of the later discussion that, following the rigors of cutting thin sections of hairs for the TEM, one often finds that splits have developed in the CMC along the line of the upper [B-layer but rarely in the lower [B-layer (34). This highlights the possibility of specific mechanical weakness associated with the upper [B-layer. On the other hand, it remains unresolved by TEM observation whether such failure occurs cohesively within the bulk of the upper [B-layer, or adhesively at one or other of its interfaces with the A-layer or with the central 8-layer. COVALENTLY LINKED FATTY ACIDS IN THE CUTICULAR B-LAYERS At a point in time when scientists thought they knew most of the basic chemistry of mammalian keratin fibers, the remarkable discovery was made of fatty acids covalently attached to the outer surfaces of wool fibers (35) and contained within the bulk of almost all mammalian keratin fibers (36,37), apart from the lowest order of Mammalia, the monotremes (38). What followed was a flurry of research activity, led principally by the Australians, toward understanding the detailed chemistry, modes of attachment, and location of these fatty acids. The excellent up-to-date review on the subject by Jones and Rivett (1) is a fine tribute to the memory of the late Don Rivett and the part he played in this pioneering work. Selected information is now drawn from this review for further discussion.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)