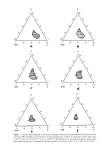
HUMAN HAIR CUTICLE 33 100- 80- 60- 40- i -20- Normal-acids ca. 25 C ø atn=21 Iso-acids Anteiso-acids I ' , i , ' , ill liii,i ,i,l l i 10 20 30 4046 No. carbon atoms for CaHz. Oz acids Figure 6. Graph showing how the melting points of saturated fatty acids vary with carbon content, according to whether they are unbranched (normal-) or contain a methyl branch in the iso- or anteiso- configurations. Note that the melting points of the normal- and iso-fatty acids are similar but that, for a given number of carbon atoms, the melting point for the anteiso-acids is at a much Dower temperature. Taken from Weitkamp (46). methyl group to rotate along the locus of a cone (with the kink at the apex). Such increases in segmental volume and disorder will be important entropic determinants for the liquid-like behavior of the chain terminus of 18-MEA. For one end of 18-MEA to be fixed rigidly and the other end to be endowed with high segmental mobility would seem to belie functional purpose. On the other hand, it is not clear why nature has chosen a chain of such a specific length or, if terminal mobility was required, why this was not accomplished by unsaturation, except that this would have made the lipids more susceptible to oxidative degradation (49). In the context of degradation, K6rner et al. (50) have shown that exposure of wool fabric to artificial sunlight significantly reduces its 18-MEA content and have suggested that the tertiary carbon at position 18 might be susceptible to hydroperoxidation. Such adducts have not

34 JOURNAL OF COSMETIC SCIENCE been identified among the other fatty acids during chemical analysis, but it remains an attractive possibility that this type of degradation might account for the decrease in hydrophobicity of the hair's surface in going from root to tip (45). CONFORMATION OF LIPIDS IN THE [•-LAYERS It would be useful to know accurately the thickness of the cuticular lB-layers as a starting point for considering the conformational disposition of the lipids within them. From TEM observations of transversely sectioned hairs, various authors have quoted the thick- ness of the lB-layers of the CMC as being somewhere between 2.5 and 4.0 nm. This is unfortunate because the lower value would be consistent with a monolayer of an ex- tended lipid chain such as 18-MEA, whereas the higher value is what might be expected for a typical lipid bilayer. One should not underestimate the difficulty of using the TEM to measure the thickness. Even assuming accurate calibration of the microscope at the high image magnifications required for the job, the main problem is that the lB-layer is contained within a section of the hair whose thickness is approximately 20 times that of the lB-layer. Even small deviations in planarity of the lB-layers within the section, or small angles of tilt in the plane of the lB-layer from being parallel to the electron beam, will serve to confound the relevance of the measurements. The problem might be resolved by using a TEM equipped with a stage goniometer to tilt the sections incre- mentally about the plane of the lB-layer, but such concerted studies have not been reported. As indicated above, great uncertainty exists as to whether the lipids in individual lB-layers of the CMC are monolayers or bilayers. Current evidence points to a monomo- lecular layer of 18-MEA being attached to the A-layer of each cuticle that constitutes the whole or part of the upper lB-layer. One presumes that the other covalently linked fatty acids, which occur in cuticle in amounts approximately equal to that of the 18-MEA, are attached to the inner layer and constitute the whole or part of the lower lB-layer. Stoichiometrically this accounts for all the covalently linked fatty acid normally present in the cuticle, and so one must exclude the possibility of there being further monolayers of covalently linked fatty acid attached to the surfaces of the 8-layer that might other- wise form bilayers with the existing covalently attached lipids in the upper and lower lB-layers. What remains unresolved are the types, amounts, and disposition of free lipids within the cuticular CMC. Some of these might be located within the body of the 8-layer, but then the ease with which this component can be stained with heavy metal compounds would seem to belie a greater level of hydrophilicity than would be the case for a structure containing significant amounts of lipid. More likely it is that these other lipids form pseudoclassic bilayers with the covalently bound lipids of the upper and lower lB-layers, the polar ends of these free lipids interfacing with the surfaces of the 8-layer. More research will be required to test the veracity of such a model. Uncertainty also exists about the thickness the upper lB-layer presents at the outer surface of degreased fibers. From the TEM of transverse fiber sections, thicknesses of from 2.5 nm (33) to 6.0 nm (1) have been quoted. To add yet further to this uncertainty, Ward et •/. (8) determined by X-ray photoelectron spectroscopy that the thickness of the hydrocarbon layer afforded by 18-MEA at the surface of wool fibers is only a matter of 0.9 nm. Zahn et •l. (18) presented a model in keeping with this latter thickness, in
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)