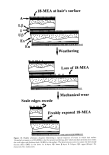
HUMAN HAIR CUTICLE 35 which surface chains of 18-MEA were folded back on themselves. This folded-chain model might be conformationally adopted as a mean for burying the intense hydropho- bic anteiso terminus of 18-MEA when solvent-extracted fibers are exposed to water. On the other hand, one must not lose sight of the fact that animal hairs are normally covered with sebaceous excretions where the 18-MEA chain can be expected to be expected to intercalate with those lipid chains. Indeed, the highly mobile anteiso terminus of 18- MEA could under such circumstances facilitate the spreading of these extraneous lipids across the fiber surface by reducing interfacial shear strength. Considerable controversy thus currently exists as to the actual thickness of the surface [3-layers and the attendant conformation of 18-MEA. This author believes these uncertainties might be resolved by concerted atomic force microscope (AFM) investigations of the surfaces of solvent- extracted hairs. He envisages that the thickness of the external 18-MEA layer might be determined from force-distance plots for the vertical penetration of the surface lipid by a sharply profiled AFM probe tip, where the lower end of the lipid chains would be sensed by a rapid rise in resistance as the crosslinked proteinaceous supporting layer is reached. He also anticipates that by AFM it will be possible to gain considerable insight into the molecular rheology of the surface 18-MEA by making use of a transversely oscillating probe tip. THE PROTEIN SUPPORTING 18-MEA Current evidence is that 18-MEA at the cuticle surface is attached via thioester linkages to the cysteine residues of an underlying layer of protein. The connecting cysteine residues would need to be presented to the outer surface of this protein in sufficient density to maintain rnonornolecular coverage of the 18-MEA. One therefore expects these cysteines to be in pseudohexagonal array and presented at an average center-to- center separation of the order of 0.936 nm to satisfy the monolayer condition (N.B.: 0.936 nm is the diameter consistent with the end-on area for each 18-MEA molecule-- cf. Appendix 1). Negri et •l. (51) have already suggested that the protein could perhaps be in the form of a [3-pleated sheet in which extended protein chains ([3-strands) lie parallel and hydrogen-bonded to each other. Such proteins offer the opportunity for amino-acid side chains to be presented at one side of the sheet at intervals of between 0.64 and 0.68 nm along each strand, for strands 0.48 nm apart. The existence of amino- acid side chains on the other side of the sheets at the same density means that the protein wouldn't need to possess a particularly high cysteine content to satisfy bonding with 18-MEA. On the other hand, a high cysteine content would be expected to satisfy the needs for intermolecular disulphide bonding on the obverse side of the sheet (i.e., between adjacent [3-strands within the sheet and with other underlying cysteine-rich proteins). Attempts to isolate and analyze fragments of the proteins still attached to 18-MEA have so far proved unsuccessful (45,50). This is unfortunate because such an approach would provide the potential for identifying the corresponding gene and for obtaining a complete amino acid sequence for the protein to which 18-MEA is attached. DIFFERENTIAL FRICTION AND ITS ORIGIN A property of human head hairs, as for all undamaged mammalian hair fibers, is that the coefficient of friction for rubbing their surfaces in a root-wise direction is significantly

36 JOURNAL OF COSMETIC SCIENCE greater than for rubbing in a tipwise direction. This directional friction effect (or DFE) can be readily demonstrated by gently rubbing a hair between the thumb and forefinger, whereupon the hair will move towards contact with its tip, an effect first reported by Monge in 1790 (52). The DFE is a principal determinant for tangling in arrays ofkeratin fibers (see below). A convenient descriptor, which accords well with propensity for tangling (or felting shrinkage in the case of wool fabrics), is the difference between the two frictional coefficients as a proportion of the sum of the two coefficients (53): DFE = (p• - pw)/(p• + Pw) (Eq. 1) where p• and p• are, respectively, the against-scales and with-scales coefficients for friction. There has never been any doubt that the main reason for directional differences in friction in mammalian keratin fibers lies with the architectural asymmetry of their surfaces, viz., of cuticular scales overlapping like tiles on a roof. What has been uncer- tain, however, are the underlying microscopic and molecular processes of frictional interaction with the fiber surfaces, and how these are influenced by topographic asym- metry. Many different methods have been used to measure frictional interaction between hair fibers and to demonstrate DFE (54-56), but meaningful analysis of the results for modeling frictional processes, as they might relate to realistic interactions between hairs on the head or during the felting of wool, undoubtedly have been confounded by the use of high experimental loading levels. Significant understanding of friction in human hair, however, has been provided by the elegant work of Adams eta/. (57), who investigated the forces of interaction in the motion of a lightly tensioned hair at right angles across a cantilevered counter fiber, where the type and diameter of the counter fiber was varied. They concluded that friction in hair does not involve major mechanical engagement or a ploughing process such as against the scale edges, as had previously been suggested (54). Convincing evidence was presented that the directional friction effect arises ac- cording to the concepts of an adhesion model where interface shear frictional processes occur in different geometric configurations according to the direction of motion. The layer of 18-MEA at the surface of undamaged hair undoubtedly has a critical influence on its frictional behavior, the high molecular mobility of the outwardly presented anteiso-configuration serving to maintain a low free surface energy and a low interfacial shear strength in frictional contacts with other surfaces and with other hairs. When 18-MEA is specifically cleaved from the surface of wool with anhydrous potas- sium tertiary butoxide in tertiary butanol, the surfaces become anionic and hydrophilic and there is a dramatic increase in friction (58). Despite this, the fibers continue to exhibit a DFE, as is reflected by the continuing capacity of the wool fabric to undergo felting shrinkage, albeit much less than in the untreated fabric (59). Examination of the butoxide-treated wool fibers in the SEM showed there was little or no change to the fiber's pattern of overlapping scales (59). What was particularly significant about Leeder and Rippon's (59) work was their demonstration that felting shrinkage, interfiber fric- tion, and wettability of the butoxide-treated wool were almost completely restored to those of the untreated wool by further treatment with a cationic-substituted stearamide. By these processes, the covalently bound 18-MEA at the surface of the original wool had been effectively replaced by an alternative fatty chain, now attached by ionic interaction to the fiber surface. In this regard, there seems little doubt that the advantage of cationic
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)