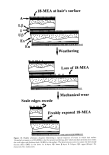
HUMAN HAIR CUTICLE 43 in the TEM, it is noticeable that portions of the CMC containing the 8-layer are located much more frequently on the endocuticular side of each fragment than on the side with the A-layer. This observation, taken with that of splitting seen in hair sections (noted above), points to the upper [•-layer as a plane of weakness and of preferred separation during dry combing. As to precisely where at the molecular level the split propagates, this is likely to be at the surface of the terminal anteiso-groups of the 18-MEA where the shear strength is predictably low. During the wet combing of hair, the softened endocuticle is a preferred plane for fracture, but even in this case, the granular endo- cuticular debris is very quickly eliminated in normal hair with shampooing and dry handling (65), again with a preference for revealing a fresh surface of 18-MEA. Combing of otherwise undamaged hair yields fragments the largest of which are platelets 1 to 2 lnm across. They are angular in shape and their edges are more or less perpen- dicular to each sheet, characteristics that seem consistent with brittle fracture, as the surface scale edges of the original fibers have been bent backwards. This is surprising, given the general traverse of the comb from root to tip in a "with-scales" direction. One presumes, therefore, that the local orientation of fibers in the array on rare occasions has conspired in against-scales frictional contacts, either between the tines of the comb and the hair or, more likely, between the hairs themselves, to produce the platelet-like fragments. The chipping away of scale edges on hair and the exposure of fresh 18-MEA at the hair surface is normally sufficiently gradual to sustain the hair for most of its lifetime, albeit the number of overlying cuticle cells also gradually diminishes from the root to the tip of the fiber (4). Indeed it is pertinent to mention that, since the average scale overlap distance is 5 lnm and the scale measure approximately 60 lnm in a longi- tudinal direction, there is the opportunity for complete restoration of 18-MEA at the hair surface twelve times before the cuticle is completely eliminated. The rate of loss of cuticle in the hairs of any person is determined by several factors, which include extent of exposure to sunlight and cosmetic habits. The layer of 18-MEA seems to have been conspired as a weak point in the upper I•-layer to facilitate preservation of the DFE, but then its interfacial strength is sufficiently high to maintain the structural cohesiveness of the cuticle as a whole. Even more remarkable is the fact that its strength is sufficiently high, coupled with a subtly balanced bending shear strength in the single thickness of a cuticle cell, to ensure that only small pieces mechanically fracture from the surface scale edges. SUMMARY AND CONCLUSIONS The hair cuticle presents: 1. Tough outer layers (A-layer and exocuticle) to help withstand chemical insult and an associated sublayer (the endocuticle) that helps the hair to withstand frictional and impactive insults. 2. Limited bending stress to brittle fracture, ensuring that only small pieces of cuticle are shed by mechanical abrasion. 3. A layer of 18-MEA located in the upper I•-layer of the CMC that is responsible for the low surface energy and low coefficient of with-scales friction of the hair's outer surface. 4. Low with-scales friction that, in association with the high against-scales friction, maintains directionally dependent frictional behavior.

44 JOURNAL OF COSMETIC SCIENCE 5. Differential friction that makes the hair on the head naturally disentangling and aligned and aids in the ejection of scalp surface detritus. 6. An upper [3-layer that, by its liquid-like behavior and relative mechanical weakness, ensures that fresh 18-MEA is presented to the hair's outer surface as mechanical attrition of the cuticle takes place. This aids in the preservation of differential friction and the associated advantages. APPENDIX 1 NUMBER OF LAYERS OF 18-MEA IN THE CUTICLE Here we compare Valldorfs (45) analyses for the 18-MEA content of root-end hair with theoretical estimates and derive an estimate of the number of layers of 18-MEA in the cuticle. Our initial assumptions are that the European hairs Valldorf used correspond to circular rods of 70 t•m average diameter (66) and that the root-end of each hair has ten cuticle cell layers at its periphery. If we consider initially that the entire outer surface of each hair is covered with a monolayer of 18-MEA, then the mass of 18-MEA in a given length of the fiber will be given by (surface area of hair) x (moleclar weight for 18-MEA) (surface area of each 18-MEA molecule) x (Avagadro's number) The surface area of the hair can be readily calculated if we know its diameter (70 pm) and length (for initial convenience we will choose 10 mm). The molecular weight of 18-MEA is 326, the surface area occupied by the anteiso-terminus of each 18-MEA molecule is 68.89 x 10 -2ø m 2 (45), and Avagadro's number (i.e., the number of molecules in one gram mole) is 6.025 x 1023. Upon substituting these quantities in the equation, the theoretical amount of 18-MEA covering the outer surface of a 10-mm length of the hair will be 1.728 x 10 -9 g. The mass density of hair is normally accepted to be 1.30 g cm -3, and on this basis, the mass ofa 10-mm length of 70-pm hair is almost exactly 50 pg. A monolayer of 18-MEA therefore corresponds to 34.6 pg of 18-MEA per gram of hair. Linear regression of Valldorf's (45) practical analyses for the amount of 18-MEA with distance along a hair sample yields a figure of 407.9 pg of 18-MEA per gram of the hair's root end. This corresponds to 11.8 theoretical monolayers of 18-MEA in the hair cuticle. Given our normal expectation of ten layers of cuticle at the root-end of human hair, this is in reasonable accord with a total of eleven monolayers of 18-MEA comprising one layer on the outer-facing surface of each cuticle cell, one monolayer in each of the nine intercuticular cell membrane complexes, and a further layer at the surface of the cortex. Interestingly, the results of Negri et al, (51) for the 18-MEA content of wool (2.3 theoretical layers) are consistent with this general model. Negri et al.'s original results were interpreted on the basis of a monolayer of 18-MEA around the entire perimeter of each cuticle cell. At any given point along a wool fiber there are either one or two layers of cuticle as provided by a single layer of scales overlapping for an average of approximately 0.3 of their length. The 2.3 theoretical layers can thus be alternatively ascribed to one monolayer at the surface of the fiber, one monolayer at the surface of the
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)