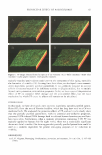
14 � ... 6 5 4 (,) ,, :e- 2 ..J 0 Cer TAURINE AND PREVENTION OF DRY SKIN Chol FA CS04 □ tadays ■ t5days O mM Ta urine ■ t5days 0.3 mM Taurine CE PC PE 7 Figure 4. Development of barrier lipid levels in the presence of taurine. Individual lipid levels were quantified in reconstructed epidermis after 17 and 22 days of culture in the absence of taurine. Incubation in the presence of 0.3 mM taurine in the culture medium was performed between days 17 and 22. Data represent the mean +/- SD (n=4). Cer, ceramide Chol, cholesterol FA, free fatty acids CS04, cholesterol sulfate CE, cholesterol esters PC, phosphatidylcholine PE, phosphatidylethanolamine. *p 0.05 **p 0.01. _- -� .:.. - ·- . . ' = .. . � -;- ---� -- ! fl.;_. Figure 5. Morphology of reconstructed epidermis after incubation with taurine. Cultures were incubated for five days in the absence (A) or presence (B) of 0.3 mM taurine, and harvested 22 days after airlift. Magnification 200 x. Together, these data demonstrate that the effect of taurine on epidermal lipid metabo- lism is restricted to the class of barrier lipids. Epidermal lipid synthesis is regulated by changes in the permeability barrier function. Barrier perturbation stimulates the synthesis of all three lipid classes (ceramides, cho- lesterol, and fatty acids) required for barrier formation (14,26,27). Studies by Man et al. (28) revealed that the correct molar ratio of those three lipid classes (i.e., approx. 1:1:1) is of critical importance for barrier integrity. Therefore, it is important to note that taurine stimulated the synthesis of ceramides, cholesterol, and fatty acids to a similar extent. Because taurine is present in normal human skin, it might also represent an important standard ingredient for keratinocyte cell culture media to optimize the barrier competence of epidermal in vitro models. The molecular mechanism of taurine-mediated lipid synthesis and barrier repair remains unknown. Recently, it has been shown that IL-lalpha stimulates barrier recovery in aged mice as well as epidermal lipid synthesis in cultured human keratinocytes (29). Because taurine impairs the SDS-induced stimulation of 11-lalpha, it is evident that the effect of taurine on barrier repair is not mediated by 11-lalpha. However, IL-1 receptor-

8 JOURNAL OF COSMETIC SCIENCE deficient mice display normal barrier repair kinetics, indicating that signalling pathways independent from IL 1-alpha predominate the barrier repair response, including stimu- lation of lipid synthesis (30). Taurine inhibits the activity of potassium ion channels (31). It has been shown that potassium ions inhibit permeability barrier repair (32). Therefore, taurine might regu- late barrier function by altering the intraepidermal transport of potassium ions. It has been shown that glycerol accelerates barrier recovery after acute disruption (33). However, the effect of taurine appears to be much more prominent because in this study a significant effect was observed at 1 % taurine in the presence of 2.5% glycerol. In contrast, the vehicle containing 2.5% glycerol alone had no significant effect on SDS- induced barrier perturbation. UV irradiation also triggers skin inflammation, barrier lipid synthesis, and taurine accumulation (7 ,34}. Therefore, future studies will focus on the effects of taurine on UV-induced skin irritation. CONCLUSION Our data suggest a role for taurine in preventing surfactant-induced dry and scaly skin by modulating the proinflammato ry response and stimulating epidermal lipid synthesis. Because barrier repair is diminished in aged epidermis (3 5 ), taurine might be especially useful to prevent surfactant-induced dry and scaly skin in the elderly. ACKNOWLEDGMENTS We thank Mrs. Christiane Bossmann-Tesch and Dr. Daniela Kessler-Becker from Hen- kel KGaA (Duesseldorf, Germany) for their excellent support with skin lipid analysis and in vitro skin irritation. We also thank Dr. Vera Meienschein, Dr. Thomas Gassen- meier, Dr. Ruediger Graf, and Dr. Michael Kock from Phenion GmbH & Co KG (Frankfurt, Germany) for their helpful discussions. REFERENCES (1) R.R. Warner, M. C. Myers, and D. A. Taylor, Electron probe analysis of human skin: Determination of the water concentration profile,]. Invest. Dermatol., 90, 218-224 (1988). (2) P. J. Caspers, G. W. Lucassen, E. A. Carter, H. A. Bruining, and G. J. Puppels, In vivo confocal Raman microspectroscopy of the skin: Noninvasive determination of molecular concentration profiles,]. Invest. Dermatol., 116, 434-442 (2001). (3) M. B. Burg, Molecular basis of osmotic regulation, Am. J. Physiol., 268, 983-996 (1995). (4) F. X. Beck, A. Burger-Kentischer, and E. Muller, Cellular response to osmotic stress in the renal medulla, Pflugers Arch., 436, 814-827 (1998). (5) M. V. Lobo, F. J. Alonso, A. Latorre, and R. Martin del Rio, Taurine levels and localisation in the stratified squamous epithelia, Histochem. Cell Biol., 115, 341-347 (2001). (6) G. Janeke, W. Siefken, S. Carstensen, G. Springmann, 0. Bleck, H. Steinhart, P. Hoger, K. P. Wittern, H. Wenck, F. Stab, G. Sauermann, V. Schreiner, and T. Doering, Role of taurine accumu- lation in keratinocyte hydration,]. Invest. Dermatol., 121, 354-361 (2003). (7) U. Warskulat, A. Reinen, S. Grether-Beck, J. Krutmann, and D. Haussinger, The osmolyte strategy
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)