6 JOURNAL OF COSMETIC SCIENCE 45 �--------------------� 40 35 � 30 I! :S 25 � 20 � 15 10 5 0 1. SDS 2. untreated p0.05 c-7 1. SDS 2. Vehicle 1.SDS 2. Vehicle+ 1% Taurine Figure 3. Effect of taurine on skin barrier function to water loss challenged by repeated exposure to SDS. Skin barrier function was repetitively perturbed by occlusive application of 0.75% SDS for 30 min twice daily for a total of five days. After removal of occlusive patches, the skin sites were either left open or treated with the vehicle or the vehicle containing 1 % taurine. The rate of transepidermal water loss (TEWL) was measured immediately before each application of SDS. All data were normalized to the starting value, and an integrated TEWL was calculated including the data from a total of ten measurements. Data represent the mean +/- SEM (n= 18). known whether taurine also affects epidermal barrier integrity in the absence of an exogenous challenge. It is interesting to note that taurine regulates osmotic water transport in renal inner medullary cells (3,4). Together with recently published evidence for an epidermal tau- rine transporter (6,7) our data indicate that taurine has similar functions in the kidney and the skin. Skin barrier homeostasis requires several metabolic steps culminating in the synthesis and secretion of barrier lipids (i.e., ceramides, cholesterol, and free fatty acids) by the outermost granular keratinocytes (13,14,25). Exposure to a dry environment increases keratinocyte uptake of taurine (6) and barrier lipid deposition to the stratum corneum (12). Because taurine significantly improves skin barrier function in vivo, taurine accu- mulation might trigger barrier lipid synthesis. To test this hypothesis, reconstructed epidermis was incubated in the presence of 0.3 mM taurine, and epidermal lipids were extracted and quantified after five days. Taurine was supplemented to the culture medium because topical application of an emulsion results in a contamination of epi- dermal lipids with various exogenous esters of fatty acids that co-migrated with specific ceramide fractions on thin-layer chromatography (data not shown). With the exception of a decrease in colesterol sulfate levels (p 0.01), lipid levels remained unchanged between day 0 and day 5 in the absence of taurine (Figure 4). In contrast, incubation of reconstructed epidermis with 0.3 mM taurine for five days significantly increased the levels of all three classes of barrier lipids (ceramide, cholesterol, and fatty acids) when compared to the untreated control (Figure 4). Increased epidermal availability of cho- lesterol also extended the pool of cholesterol sulfate and cholesterol esters. In contrast, the levels of phosphatidylcholine and phosphatidylethanolamine did not change signifi- cantly (Figure 4). In the epidermis, phospholipids are restricted to the viable cell layers. In accordance with the lipid results, the number of viable cell layers did not change after incubation of reconstructed epidermis with 0.3 mM taurine for five days (Figure 5).

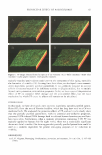
14 � ... 6 5 4 (,) ,, :e- 2 ..J 0 Cer TAURINE AND PREVENTION OF DRY SKIN Chol FA CS04 □ tadays ■ t5days O mM Ta urine ■ t5days 0.3 mM Taurine CE PC PE 7 Figure 4. Development of barrier lipid levels in the presence of taurine. Individual lipid levels were quantified in reconstructed epidermis after 17 and 22 days of culture in the absence of taurine. Incubation in the presence of 0.3 mM taurine in the culture medium was performed between days 17 and 22. Data represent the mean +/- SD (n=4). Cer, ceramide Chol, cholesterol FA, free fatty acids CS04, cholesterol sulfate CE, cholesterol esters PC, phosphatidylcholine PE, phosphatidylethanolamine. *p 0.05 **p 0.01. _- -� .:.. - ·- . . ' = .. . � -;- ---� -- ! fl.;_. Figure 5. Morphology of reconstructed epidermis after incubation with taurine. Cultures were incubated for five days in the absence (A) or presence (B) of 0.3 mM taurine, and harvested 22 days after airlift. Magnification 200 x. Together, these data demonstrate that the effect of taurine on epidermal lipid metabo- lism is restricted to the class of barrier lipids. Epidermal lipid synthesis is regulated by changes in the permeability barrier function. Barrier perturbation stimulates the synthesis of all three lipid classes (ceramides, cho- lesterol, and fatty acids) required for barrier formation (14,26,27). Studies by Man et al. (28) revealed that the correct molar ratio of those three lipid classes (i.e., approx. 1:1:1) is of critical importance for barrier integrity. Therefore, it is important to note that taurine stimulated the synthesis of ceramides, cholesterol, and fatty acids to a similar extent. Because taurine is present in normal human skin, it might also represent an important standard ingredient for keratinocyte cell culture media to optimize the barrier competence of epidermal in vitro models. The molecular mechanism of taurine-mediated lipid synthesis and barrier repair remains unknown. Recently, it has been shown that IL-lalpha stimulates barrier recovery in aged mice as well as epidermal lipid synthesis in cultured human keratinocytes (29). Because taurine impairs the SDS-induced stimulation of 11-lalpha, it is evident that the effect of taurine on barrier repair is not mediated by 11-lalpha. However, IL-1 receptor-
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)