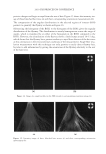
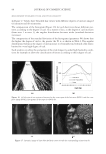
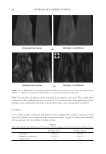
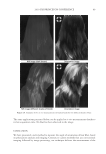
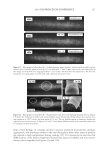
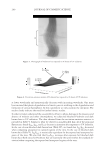
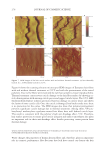
2010 TRI/PRINCETON CONFERENCE 199 PHOTODAMAGE OF HAIR As part of the integument, the outermost organ of the body, hair is constantly exposed to solar radiation. Normally, UVB radiation (290–320 nm) is of great concern due to it deleterious effects in the skin resulting in DNA damage, possibly leading to carcinomas or melanoma. Historically, UVA radiation (320-400 nm) was thought to be less signifi - cant since its light waves are much lower in energy than UVB radiation. In the last de- cade, more emphasis has been placed on the recognition that UVA is also damaging (greater quantities reach the Earth’s surface) and causes a number of free radical reactions that lead to lipid peroxidation, protein degradation, and even, DNA damage in skin. In hair, lipid peroxidation, can result in altered surface properties and structural organiza- tion while protein degradation affects the overal morphological integrity of the fi ber. As already noted, measurement of Trp by spectrofl uorescence was employed to monitor the health state of hair exposed to solar radiation (10). In the current study, we utilize sim- ilar methodology however, we generate excitation-emission matrices in order to probe the behavior of all fl uorophores present in hair. Figure 5 contains a photograph of a hair tress exposed to 96 hours of UV radiation. As noted in the fi gure, only the middle section of the tress was exposed, resulting in photobleaching of the yellow pigment naturally present in Piedmont hair. An excitation-emission matrix for the photo-exposed Piedmont hair is presented in Figure 6. Similar to what we observed in bleached hair, photoexposure reduces the peak intensity for the bands at longer excitation wavelengths. In untreated Piedmont hair, there is convergence of numerous bands starting from the principal kynurenine peak (λex = 394 nm, λem = 465 nm) to the longest excitation wavelength (λex = 450 nm, λem = 509 nm). After photo-exposure, the bands at longer wavelengths decrease relative to the principal kynurenine band. This same effect was observed in bleached hair (see Figure 3) and coincides with a loss of the natural yellow hue of Piedmont hair. A summary of the data from excitation-emission matrices for dark brown and Piedmont hair is provided in Table IV. For all of the monitored fl uorophore bands (ITrp, IKyn, and I509), we see a decrease in intensity when comparing the unexposed to exposed regions. In the case of dark brown hair, ITrp/IKyn increases in the region of the tress exposed to UV light whereas in Piedmont hair similar readings are obtained for both regions. At fi rst glance, one may speculate that this is a melanin-associated phenomenon—the kynure- nines are protected more than Trp by melanin in dark brown hair. In terms of absolute absorbance, we would expect the contrary to be true since melanin absorbance is greater Table III Peak Intensity Values and Pertinent Peak Ratios for Dark Brown and Piedmont Hair Exposed to Thermal Treatment ITrp IKyn I509 ITrp/IKyn I509/IKyn Dark brown 24,550 ± 2,616 103,050 ± 5,586 33,250 ± 7,283 0.239 ± 0.038 0.321 ± 0.053 Dark brown- thermal 15,100 ± 849 108,000 ± 4,243 29,650 ± 4,172 0.140 ± 0.002 0.276 ± 0.049 Piedmont 44,600 ± 5,657 2,030,000 ± 98,995 1,275,000 ± 63,640 0.022 ± 0.001 0.621 ± 0.062 Piedmont- thermal 18,900 ± 1,838 1,800,000 ± 127,279 1,540,000 ± 28,284 0.010 ± 0.000 0.858 ± 0.076 Data are provided for the unexposed and exposed regions of the hair tress.

JOURNAL OF COSMETIC SCIENCE 200 at lower wavelenths and monotonically decreases with increasing wavelenth. One must bear in mind that photo-degradation is a kinetic process resulting in the degradation and formation of various fl uorophores. In this experiment we only examine the end point. The results clearly indicate the need for further kinetic studies. In order to better understand the susceptibility of hair to photo-damage in the presence and absence of melanin and other chromophores, we subjected bleached Piedmont and dark brown hair to UV radiation. The data obtained from the excitation-emission matrices is provided in Table V. Similar to what we observe in non-bleached hair, all of the principal fl uorescence bands (ITrp, IKyn, and I509) decrease in intensity after exposure to UV radiation. In the case of non-bleached dark brown hair (Table IV), we found that ITrp/IKyn decreased when comparing unexposed to exposed regions of the tress. In the case of bleached dark brown hair (Table V), ITrp/IKyn is statistically equivalent for the exposed and unexposed re- gions of the tress. We also fi nd that I509/IKyn increases after exposure for bleached dark brown hair. Both ratios for Piedmont hair are essentially the same (within the standard de- viation) when comparing the exposed and unexposed regions of the tress. Figure 5. Photograph of Piedmont hair exposed to 96 hours of UV radiation. Figure 6. Excitation-emission matrix of Piedmont hair exposed to 96 hours of UV radiation.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)