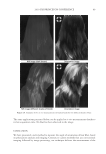
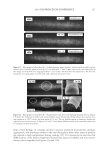
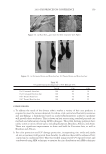
JOURNAL OF COSMETIC SCIENCE 232 phosphate (pH 7.8) - 20 mM DTE and then pulverized with a magnetic stirring bar for several hours. Aliquots of the solubilized and insoluble fractions were alkylated with iodo- acetamide, and the solubilized protein was recovered as a fl occulent precipitate by addi- tion of ethanol to 70%. The two fractions were rinsed in 70% ethanol and fresh 0.1 M ammonium bicarbonate and then digested with stabilized trypsin (≈1% by weight) in the bicarbonate buffer adjusted to 10% in acetonitrile. For this purpose, TPCK-treated bo- vine trypsin was stabilized by reductive methylation (17). Yields were improved by diges- tion for 2-3 days at room temperature with daily additions of trypsin. At the conclusion of digestion, the samples were clarifi ed, taken to dryness, submitted for mass spectromet- ric analysis and the data presented using Scaffold Proteomics Software (18). The reports permit ready access to many details of the protein identifi cation including degree of pep- tide coverage, spectral characteristics, unique peptides and statistical description. The original fi ndings from our analyses (19) were that the solubilized protein fraction of hair shaft was comprised of a score of keratin and keratin-associated proteins known to be present, while the insoluble fraction was much more complex. Using two chromatographic steps of peptide separation permitted identifi cation of some 350 proteins in the latter fraction originating from all regions of the cell. Careful examination of the peptide frag- mentation patterns permitted identifi cation of methylated lysines in a dozen proteins and evidence of ubiquitination as a glycylglycine posttranslational lysine modifi cation of ubiq- uitin itself. For these analyses, the peptide digest was fractionated by ion exchange chro- matography, and each fraction was then submitted to online reverse phase chromatography and mass spectrometry (LC-MS/MS). In the interest of higher throughput in later work, the fi rst step of ion exchange chromatography was omitted subsequently. Although not as comprehensive, the results typically identify 100 proteins in a given sample, including novel components, a benefi t of this discovery approach. Quantitating protein amounts in the shotgun approach is a challenge in general due to the unknown yields of peptides in mass spectrometry. Contributing further to the uncertainty are the possibly incomplete yields from digestion of cross-linked material and the inability to identify peptides derived from protein cross-linkage sites. Since only a minority of ly- sines are involved in isopeptide linkages, estimated as ≈18% for epidermal callus (20), suffi cient peptides are generated from the proteins to permit successful identifi cation. With these caveats, estimates have been made of relative amounts based on the empirical relation between the protein abundance and the number of unique peptides (i.e., not counting overlaps in sequence coverage) detected compared to those predicted (21). Although only semi-quantitative, estimates derived by this relation, called an exponen- tially modifi ed protein abundance index, offer a convenient way to rank proteins in relative abundance (22). APPLICATIONS OF SHOTGUN PROTEOMICS This approach to human hair syndromes exhibiting structural abnormalities plausibly will permit identifying protein defi ciencies, although it is not guaranteed to do so. In the case of a family displaying autosomal recessive woolly hair, samples from affl icted offspring did not show any defi ciencies in the prominent proteins identifi ed (23). In this instance, the lipase H gene was found to be defective by genetic testing. The availability of numerous mouse strains with anomalous structures suggests proteomic analysis could be useful in

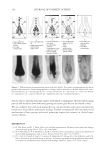
2010 TRI/PRINCETON CONFERENCE 233 characterizing bases for the defects. AKR mice, for example, display hair with an “interior defect” consisting of haphazard patterns of medulla cell arrangement due to mutations in steroyl O-acetyltransferase 1 (24). In strains without this defect, where medulla cells are typically in an orderly alignment with regular spacing, electron microscopy of mature hair shafts reveals projections from the cortex into the middle of each medulla cell. The AKR strain lacks these indentations, evidently permitting irregular or disorderly spacing of the medulla cells. Proteomic analysis revealed only a low level of trichohyalin, a major compo- nent of the projections, in the cortical cells of AKR mice compared to two other strains not showing this phenotype (4). In the course of this work, it was noted that all three mouse strains could be distinguished by their proteomic profi les, raising the question whether, by analogy, human hair from individuals of different ethnic origin could be distinguished by protein profi ling. This approach to analysis of hair is easily adaptable to other keratinized structures such as nail plate. The nail plate proteome resembles that of the hair shaft in its high content of keratin proteins and the more complex profi le of the insoluble fraction compared to the solubilized proteins (25). Nearly all the 30 proteins in the solubilized fraction were kera- tins and keratin-associated proteins, while the insoluble fraction was comprised of, in addition to these, cytoplasmic proteins, membrane and junctional proteins and histones (Figure 3). Many of the proteins overlapped considerably with those detected in hair shaft, notably some abundant keratins (K31, K33B, K34, K39, K85, K86), junctional proteins (DSP, JUP, DSG4, PKP1) and numerous intracellular proteins (SELENBP1, SFN, ACTB, HSPA5). Some proteins were detected only in nail plate, including certain keratins (K5, K6A, K14, K17), junctional proteins (DSG1, EPPK1) and intracellular proteins (KPRP, SERPINB12, CKB). Others seen only in hair shaft included several keratins (K40, K82) and intracelluar proteins (CTTNB1, ATP5B). The fi nding of keratins (and KPRP) found only in nail, which are also found in epidermis, supports the established view that the nail unit expresses features of both hair and epidermis (26). Figure 3. Cellular locations of identifi ed proteins in nail plate sorted into the categories keratin and keratin- associated proteins (Ker), other cytoplasmic proteins (Cyt), membrane and junctional proteins (Mem), and histones (His). Relative amounts were estimated based on unique peptides by the exponentially modifi ed protein abundance index (21,25).
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)