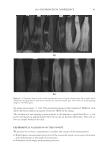
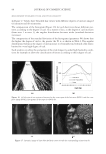
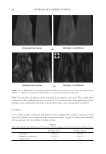
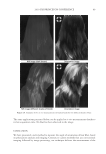
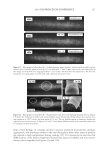
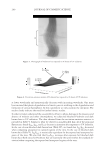
JOURNAL OF COSMETIC SCIENCE 230 great cohesiveness to the overall structure (3). The native hair shaft presents diffi culties in transmission electron microscopy due to poor penetration by embedding media without treatment to permit its diffusion into cellular interiors. A simple such treatment (Figure 1) is to incubate the hair in sodium dodecyl sulfate (SDS) under reducing conditions for an hour or two at room temperature, inducing swelling and softening of the fi ber and permitting ultrastructural visualization of bundles of intermediate fi laments in the cortex (4). More extensive detergent extraction at elevated temperature yields nearly empty cor- tical cells with their outer boundaries clearly delineated. By contrast, cuticle cells appear largely intact, with clear demarcation of the endocuticle, exocuticle and marginal band. Cells of the medulla contain remnant nuclei and amorphous deposits as well as areas with- out electron dense material. The cross-linking process in hair resembles that in the analo- gous cornifi ed appendages of bird feather and hagfi sh teeth, and thus occurs generally among vertebrates (5). Defects in the cross-linking process can be visualized microscopically in samples from epi- dermis and appendages after extraction of solubilizable protein with SDS and reducing agent. Individuals defi cient in this process can exhibit lamellar ichthyosis and related skin conditions such as congenital ichthyosiform erythroderma characterized by high rates of transepidermal water loss and prominent scaling. Instead of displaying the normally ob- served empty cells with distinct boundaries, samples of extracted epidermal scale appear fragmented and disorganized. Small pieces of nail plate show analogous alterations in the diseased state with loss of the normally obvious cell borders (6,7), while the hair cuticle shows considerably increased extraction or loss of features such as the marginal band (8,9). Structural alterations in the cortex and cuticle of uncertain basis are also seen in hair from individuals affl icted with trichothiodystrophy (8) and in the cuticle from mouse mutants such as matted and naked (10). Based on original work on wool and hair in animal models, trapping of protein constitu- ents in insoluble complexes was attributed to the action of transglutaminases found in the hair follicle (11,12), analogous to the action of factor Xllla in blood clotting (13). Studying Figure 1. Swelling of hair fi bers at room temperature. Fibers from a Caucasian male were incubated in water or 0.1 M sodium phosphate (pH 7.8), in the latter case with 20 mM DTE and/or 2% SDS. Samples were ex- amined after 6 hr except as indicated (2 hr). Shown are the means and standard deviations of diameters of 6–30 fi bers for each condition measured microscopically (16×) using Slidebook 5.0 software. Swelling and increased fl exibility are fi rst noticeable visually after 1–2 hr, at which time hair fi bers can be fi xed and embedded.

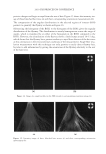
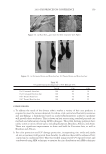
2010 TRI/PRINCETON CONFERENCE 231 the biochemical basis for the cross-linking process was assisted by the discovery that it is manifested in cultures of human epidermal cells (14). Requiring elevated calcium ions for their enzymatic activity, transglutaminases participate in the terminal stage of keratino- cyte differentiation. A major form encoded by the gene TGM1, defi cient in a large per- centage of lamellar ichthyosis cases, is located on the inside of the plasma membrane and is thereby positioned to form the cross-linked envelope at the cell periphery (15). Defects in the protein envelope are envisioned to perturb the attachment to it of the lipid barrier. Consequences of an improperly formed lipid barrier include entry of exogenous hydro- philic agents and excessive transepidermal water loss through the epidermis. Mature corneocytes consist largely of hydrophobic keratin intermediate fi laments and keratin associated proteins held together by disulfi de bonds. These components are further stabilized by proteins cross-linked to them by transglutaminase activity. In addition to the TGM1 gene product at the cell periphery, which incorporates membrane-bound and junctional proteins, a transglutaminase in the cytoplasm encoded by the TGM3 gene is available to incorporate soluble proteins (16). Our recent proteomic efforts analyzing the corneocyte have focused on these disulfi de- and isopeptide-linked components. Thus, samples of hair, nail or callus are fi rst rinsed in detergent (2%SDS) to remove soluble protein, generally negligible in amount, and adherent adventitious material to yield the sample to be analyzed. METHODOLOGICAL APPROACH Our strategy has been to separate the material solubilized in SDS dithioerythritol (DTE) from the remainder resistant to solubilization due to isopeptide bonding. To facilitate re- lease and removal of the solubilized protein, the hair was extracted fi ve times, as shown in Figure 2. Each time, it was incubated overnight at 70°C in 2% SDS - 0.1 M sodium Figure 2. Solubilized protein from human hair shaft, nail plate, or epidermis extracted in 2% SDS - 20 mM DTE - 0.1 M sodium phosphate (pH 7.8). Each extract of hair and nail consisted of daily incubation for ≈ 22 hr at 70°C and then magnetic stirring for ≈ 2 hr at room temperature. For epidermis, isolated from skin at 55°C (29), extractions were performed by heating in the above SDS - DTE - phosphate buffer solution for 5–10 min in a boiling water bath followed by vigorous vortexing. Solubilized protein was recovered by centrifugation between extractions. The slower rate of protein extraction from the nail plate refl ects the slower rate of pulveri- zation during magnetic stirring. Illustrated are the means and standard deviations of 4–6 samples of each type.
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)