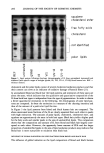
BINDING OF SURFACTANTS 195 character. Processes iii and iv are intrinsic effects of micelies to the binding equilibrium of insoluble proteins and are, thus, thermodynamic in nature. These contributions are discussed below. It has been suggested that surfactants above their CMC cause some delipidization of the stratum corneum (5,34), even though no consensus exists on this topic (35-37). An alternative view is that surfactants can remove selectively certain components such as the ceramides and in the process cause changes in the lipid composition (38). It is possible that the reasons for the discrepancies existing in the literature are essentially due to the differences in the protocols used in various studies, especially the surfactant concentra- tion and time and frequency of exposure. Our unpublished results of an ongoing TEM study of porcine stratum corneum treated with SDS for one hour, similar to the con- ditions of the binding experiments, indicate that SDS at levels above the CMC cause significant delipidization (39). Similarly, our ultrastructural studies of human skin subjected to an in vitro arm-wash protocol reported elsewhere showed that soaps caused significant lipid removal and protein damage (40). Such removal of lipids by surfactant micelies may play an important role in enhancing the diffusion of surfactant monomers and micelles to the corneocyte-matrix interface. Therefore, continued binding above the surfactant CMC is not unreasonable, even when only monomers are involved in the binding process. Note that the results of surfactant binding to delipidized corneum discussed earlier (see Figure 4) may appear to be contradictory to the proposed hypoth- esis of micelies enhancing the monomer binding by enhancing the surfactant diffusion by removing free lipids. The apparent contradiction can, however, be reconciled by noting that the delipidization study discussed earlier was done using solvents and that subsequently the skin was dried before contacting with the surfactant solution. During drying the covalently linked lipids would have formed a strong hydrophobic network that would be much more impenetrable to the surfactants than the normal corneum containing both the free lipids and the covalently bonded lipids. Yet another mechanism by which anionic micelies can influence the binding process may be by sequestering metallic ions such as calcium. Even though the role of ions such as calcium in stratum corneum cohesion is not well understood, it is known that addition of EDTA, a well-known metal ion chelator, enhances the surfactant-induced release of corneocytes (41-42). If the underlying reason for the effect of EDTA is metal ion complexation, one can expect micelies also to play a similar role and reduce the cohesion between corneocytes, which in turn can enhance the penetration of surfactants to surface regions of corneocytes. Once the surfactant molecules obtain access to protein-rich corneocyte regions, the extent of binding and especially cooperative binding in the postmicellar region is primarily determined by the inherent tendency of the surfactant monomer to bind to the proteins in the corneocyte membrane. The cooperative binding in these systems may well induce the changes in the conformation of the keratinous proteins present in the corneocyte membrane, which in turn may lead to damage to the corneum. This may require some threshold level of binding to overcome the cohesive forces arising from the conformation of the protein as well as the lipids holding the membrane together. This is similar to Breuer's concept of the "elastic energy" holding the keratinous network together (26). Surfactant binding obviously induces repulsive electrostatic forces that at some critical binding should overcome "cohesive forces" and lead to an unfolding of the protein. Thus, while all micelies, including nonionic surfactant micelies, can cause some

196 JOURNAL OF THE SOCIETY OF COSMETIC CHEMISTS removal of fluid lipids from the matrix lipid, significant damage to the protein itself will still depend upon the ability of the surfactant monomer to bind to the corneocyte membrane proteins. This is consistent with the reported observations that nonionic surfactants such as Triton X-100, in combination with some mechanical agitation, can release corneocytesrfrom the lipid matrix without damaging the membrane envelope (43). In contrast to this, treatments with harsh surfactants such as SDS or soaps result in damage to lipids and corneocyte proteins (38,39). In addition to the above mechanisms, micelles may be contributing directly to the unfolding process by creating osmotic stresses on the corneocyte envelope. It is well known that corneocytes swell when exposed to water and aqueous surfactant solutions (44). In general, the extent of corneum swelling by anionic surfactants increases with their harshness toward skin (44). It has also been reported that the extent of swelling as a function of SDS concentration attains a plateau value near the CMC of SDS (44), suggesting that monomers that bind to the corneum, including the accessible sites on the membrane, may be enhancing corneocyte swelling. This is consistent with the present observation that initial surfactant binding to the corneum exhibits a near plateau or a slowly rising region around the surfactant CMC. Interestingly, it has also been reported (44) that the surfactant-induced swelling process is reversible up to about 35 mM (1%) SDS, but is not reversible above about 70 mM (2%), indicating irreversible damage at such levels. The sharp rise in the SDS binding beyond the CMC also occurs around 60 mM, suggesting that this rise is responsible for the irreversible damage to the corneum. The origin of the sharp increase in SDS binding occurring around 60 mM may involve contributions from miceliar exclusion and the associated excluded volume effects. Be- yond the CMC, the corneocyte envelope may act as a semipermeable membrane that does not allow the micelles to penetrate into the cells. This is similar to Middleton's (45,46) concept that the corneocyte membrane acts as a semipermeable envelope holding the water-absorbing NMFs (natural moisturizing factors) within the cell and allowing water to move across the membrane. Exclusion of micelles will indeed result in an osmotic effect due to an increase in the concentration of micelles, leading to mechanical stresses on the membrane. In normal systems, one would have expected deswelling of the corneum because of osmotic stresses. Interestingly, the reported results do not indicate any deswelling above the surfactant CMC (44). This may be because the water respon- sible for the swelling is associated with the hydration of the surfactant-bound proteins and the NMFs, and the removal of the bound water may require a critical pressure (energy) that is greater than that which can be achieved by micellar exclusion. However, another pathway is available to release the osmotic stress. This may involve unfolding of the membrane proteins that are already weakened by surfactant binding, leading to a physical breakdown of the membrane structure. The unfolded proteins, in turn, expose more sites for surfactant binding. This phenomenon can also be viewed in the following manner: When surfactant binds to water-swollen proteins in a cooperative fashion, micelles are essentially transferred from solution to the "solid" protein phase. This process leads to a decrease in the chemical potential associated with the micelles. If this chemical potential difference, coupled with the free energy of cooperative surfactant- protein binding, is greater than the molecular interactions holding the protein network intact, the protein should unfold to accommodate bound micelles. Thus an increase in
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)