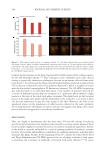
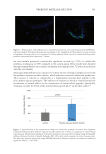
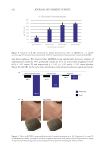
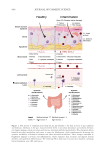
583 Specificities of Microbiota From Sensitive Skin
secretion, and facial skin surface morphology were evaluated in parallel to 16S rRNA
sequencing. No change in diversity was observed for SS and NS face skin. In the three
most common microbiota genera, including Cutibacterium, Paracoccus, and Corynebacterium,
there was no significant difference in the relative frequency among the SS and NS groups.
Nevertheless, at the genus level the authors found that in the SS group a significantly
higher frequency of rare bacteria such as Acidaminococcus, Alloiococcus, Fimbriimonas,
Pseudoclavibacter, Reyranella, and Roseococcus, but a lower frequency of Bradyrhizobium.
Spearman correlation analysis showed correlation between the microbiota genera and skin
physiological parameters. However, they found no bacterial genus with strong association
with skin physiological parameters.
A study on Caucasian women (cheek, SS evaluation using capsaicin test (61 NS 23–50
years old, 23 NS 51–69 years old, and 44 SS 20–50 years old) likewise did not show
a difference in diversity. Nevertheless, SS compared to NS with the same age class was
reported here to display a specific microbiota profile with a significant decrease in SS in
Cutibacterium, Lawsonella, and Bacillus (nonsignificant decrease for Staphylococcus), and an
increase in Corynebacerium, Snodgrassela, Kocuria, and Micrococcus.16
Finally, in 2024, Lu et al. performed a study on 10 panelists (cheek, 22–35 years old,
LAST-based recruitment) using a 2bRAD-M sequencing method enabling accurate
characterization of the low-biomass microbiome at species level. They showed in the
SS group a decrease of alpha diversity and an increase of abundance in the 3 top genera
Cutibacterium, Corynebacterium, and Staphylococcus. Among Staphylococci, S. epidermidis was
the more abundant and increased by 1.34-fold, whereas less abundant S. aureus was
the most increased (2.98-fold). They also observed that the relative abundance of rare
bacteria such as Dermabacter hominis and Chryseobacterium was significantly increased,
whereas those of Neisseria and Streptococcus decreased. Advantageously, Staphylococci
(S. epidermidis, S. capitis, and S. aureus) and Micrococcus luteus from a panelist with SS
were extracted and compared to standard S. epidermidis and aureus for their ability to
induce Interleukin-8 (IL-8) in keratinocytes. Interestingly, S. capitis and M. luteus from
the panelist with SS significantly promoted more IL-8 expression than S. epidermidis and
S. aureus.17
In this study, our goal was to first compare the composition of the skin microbiota of
individuals with NS and SS using the entire 16S rRNA gene sequencing from PacBio
technology to obtain unprecedented taxonomic resolution of the microorganisms present.
However, at the same time, we evaluated for the first time droplet-based microfluidic
technology (DBMT) to create a collection of specific bacterial species isolated from
individuals with SS and NS. DBMT was chosen as this microbiome-on-a-chip high-
throughput technology can generate more than 100 million parallel picoliter-sized droplets,
allowing the isolation of live bacterial species that grow to clonal populations at a much
higher diversity compared to standard microbiology methods when using specific culture
media. In addition, the picoliter-sized droplets containing bacteria can be isolated drop
by drop for subsequent bacterial clones’ characterization and further cosmetic ingredient
testing. The bacterial collection was used here to study the impact on bacterial growth
of cosmetic ingredients. Isolates of C. acnes from both the NS and SS collections were
also compared for their potential to stimulate the proinflammatory cytokine IL-8 in
keratinocytes with the final aim to build a screening tool using a set of bacteria previously
shown to be modulated in SS and to have impact on SS features.
secretion, and facial skin surface morphology were evaluated in parallel to 16S rRNA
sequencing. No change in diversity was observed for SS and NS face skin. In the three
most common microbiota genera, including Cutibacterium, Paracoccus, and Corynebacterium,
there was no significant difference in the relative frequency among the SS and NS groups.
Nevertheless, at the genus level the authors found that in the SS group a significantly
higher frequency of rare bacteria such as Acidaminococcus, Alloiococcus, Fimbriimonas,
Pseudoclavibacter, Reyranella, and Roseococcus, but a lower frequency of Bradyrhizobium.
Spearman correlation analysis showed correlation between the microbiota genera and skin
physiological parameters. However, they found no bacterial genus with strong association
with skin physiological parameters.
A study on Caucasian women (cheek, SS evaluation using capsaicin test (61 NS 23–50
years old, 23 NS 51–69 years old, and 44 SS 20–50 years old) likewise did not show
a difference in diversity. Nevertheless, SS compared to NS with the same age class was
reported here to display a specific microbiota profile with a significant decrease in SS in
Cutibacterium, Lawsonella, and Bacillus (nonsignificant decrease for Staphylococcus), and an
increase in Corynebacerium, Snodgrassela, Kocuria, and Micrococcus.16
Finally, in 2024, Lu et al. performed a study on 10 panelists (cheek, 22–35 years old,
LAST-based recruitment) using a 2bRAD-M sequencing method enabling accurate
characterization of the low-biomass microbiome at species level. They showed in the
SS group a decrease of alpha diversity and an increase of abundance in the 3 top genera
Cutibacterium, Corynebacterium, and Staphylococcus. Among Staphylococci, S. epidermidis was
the more abundant and increased by 1.34-fold, whereas less abundant S. aureus was
the most increased (2.98-fold). They also observed that the relative abundance of rare
bacteria such as Dermabacter hominis and Chryseobacterium was significantly increased,
whereas those of Neisseria and Streptococcus decreased. Advantageously, Staphylococci
(S. epidermidis, S. capitis, and S. aureus) and Micrococcus luteus from a panelist with SS
were extracted and compared to standard S. epidermidis and aureus for their ability to
induce Interleukin-8 (IL-8) in keratinocytes. Interestingly, S. capitis and M. luteus from
the panelist with SS significantly promoted more IL-8 expression than S. epidermidis and
S. aureus.17
In this study, our goal was to first compare the composition of the skin microbiota of
individuals with NS and SS using the entire 16S rRNA gene sequencing from PacBio
technology to obtain unprecedented taxonomic resolution of the microorganisms present.
However, at the same time, we evaluated for the first time droplet-based microfluidic
technology (DBMT) to create a collection of specific bacterial species isolated from
individuals with SS and NS. DBMT was chosen as this microbiome-on-a-chip high-
throughput technology can generate more than 100 million parallel picoliter-sized droplets,
allowing the isolation of live bacterial species that grow to clonal populations at a much
higher diversity compared to standard microbiology methods when using specific culture
media. In addition, the picoliter-sized droplets containing bacteria can be isolated drop
by drop for subsequent bacterial clones’ characterization and further cosmetic ingredient
testing. The bacterial collection was used here to study the impact on bacterial growth
of cosmetic ingredients. Isolates of C. acnes from both the NS and SS collections were
also compared for their potential to stimulate the proinflammatory cytokine IL-8 in
keratinocytes with the final aim to build a screening tool using a set of bacteria previously
shown to be modulated in SS and to have impact on SS features.