592 JOURNAL OF COSMETIC SCIENCE
Different active ingredients were tested to investigate their effect on the growth of these
species to evaluate if any would have an impact on the microbial profile. On one hand, one
of them had for example an appreciable impact on the bacterial growth of species isolated
from NS (Figure 6A, Inonotus obliquus). However, as we observed large variations for several
species of Staphylococcus and Cornybacterium, these results would need to be confirmed. On
the other hand, some ingredients had a slight influence on the growth of species (Figure 6B,
Dendrobium officinale). This type of ingredient may fit into the “microbiome-friendly” trend,
meaning an ingredient that would cause little or no damage to the NS flora, to preserve its
protective function of the skin.
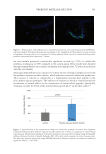
Finally, the impact of C. acnes, the most abundant bacteria found in both cohorts showing
an increase in SS was evaluated for its impact on keratinocytes. We first assessed how a
series of dilutions from 106–108 CFU/mL of living C. acnes strains derived from either
NS or SS source and placed in an insert affected the survival of keratinocytes grown
underneath. (Figure 7, left). The threshold of viability was fixed at 80% of cells viability.
The results showed no strong decrease in cell viability until 107 CFU/mL regardless of
the living C. acnes source. However, at 108 CFU/mL, all C. acnes from SS induced through
soluble factors a decrease of keratinocytes viability below the 80% threshold, with a higher
impact for SS4 and SS8 species. Then, we evaluated the production of IL-8 chemokine
by keratinocytes grown in contact with heat-killed C. acnes seeded at 106–108 CFU/mL
(Figure 7, right). IL-8 is a potent chemotactic and proinflammatory cytokine, produced
in the skin by a variety of cells in response to inflammatory stimuli, especially in SS. We
observed that at the highest dose of 108 CFU/mL, heat-killed C. acnes SS4 and SS7 are
more stressful than the C. acnes from NS or SS8 as they strongly induced IL-8 release by
keratinocytes grown in contact.
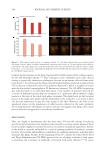
Figure 6. Illustration of growth results of 2 ingredients on the bacterial collection from normal skin. Growth
of 31 selected bacterial species evaluated with increasing concentrations of active ingredients over 24 h, 48 h,
and 72 h. A: Example of Inonotus obliquus mushroom extract at C1 to C4 (0.01%–1%), with impact on bacterial
growth. B: Example of Dendrobium officinale stem extract at C1 to C4 (0.002%–0.05%) showing minimal
changes in microbial growth. Boxes in green resulted in no significant growth difference compared to
untreated control, boxes in light gray showed growth inhibition and in dark gray growth increase. Statistics:
n =4, Dunnett’s multiple comparison test versus untreated control, significant when p 0.05.
Different active ingredients were tested to investigate their effect on the growth of these
species to evaluate if any would have an impact on the microbial profile. On one hand, one
of them had for example an appreciable impact on the bacterial growth of species isolated
from NS (Figure 6A, Inonotus obliquus). However, as we observed large variations for several
species of Staphylococcus and Cornybacterium, these results would need to be confirmed. On
the other hand, some ingredients had a slight influence on the growth of species (Figure 6B,
Dendrobium officinale). This type of ingredient may fit into the “microbiome-friendly” trend,
meaning an ingredient that would cause little or no damage to the NS flora, to preserve its
protective function of the skin.
Finally, the impact of C. acnes, the most abundant bacteria found in both cohorts showing
an increase in SS was evaluated for its impact on keratinocytes. We first assessed how a
series of dilutions from 106–108 CFU/mL of living C. acnes strains derived from either
NS or SS source and placed in an insert affected the survival of keratinocytes grown
underneath. (Figure 7, left). The threshold of viability was fixed at 80% of cells viability.
The results showed no strong decrease in cell viability until 107 CFU/mL regardless of
the living C. acnes source. However, at 108 CFU/mL, all C. acnes from SS induced through
soluble factors a decrease of keratinocytes viability below the 80% threshold, with a higher
impact for SS4 and SS8 species. Then, we evaluated the production of IL-8 chemokine
by keratinocytes grown in contact with heat-killed C. acnes seeded at 106–108 CFU/mL
(Figure 7, right). IL-8 is a potent chemotactic and proinflammatory cytokine, produced
in the skin by a variety of cells in response to inflammatory stimuli, especially in SS. We
observed that at the highest dose of 108 CFU/mL, heat-killed C. acnes SS4 and SS7 are
more stressful than the C. acnes from NS or SS8 as they strongly induced IL-8 release by
keratinocytes grown in contact.
Figure 6. Illustration of growth results of 2 ingredients on the bacterial collection from normal skin. Growth
of 31 selected bacterial species evaluated with increasing concentrations of active ingredients over 24 h, 48 h,
and 72 h. A: Example of Inonotus obliquus mushroom extract at C1 to C4 (0.01%–1%), with impact on bacterial
growth. B: Example of Dendrobium officinale stem extract at C1 to C4 (0.002%–0.05%) showing minimal
changes in microbial growth. Boxes in green resulted in no significant growth difference compared to
untreated control, boxes in light gray showed growth inhibition and in dark gray growth increase. Statistics:
n =4, Dunnett’s multiple comparison test versus untreated control, significant when p 0.05.