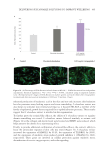
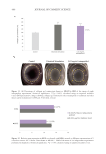
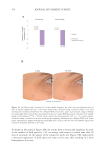
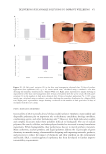
505 Evolution and Challenges of Sustainability
These AMPs help protect the skin from colonization and infection by transient
microorganisms and pathogens.
6. Commensal skin microflora may promote immunological quiescence by up-regulating
production of IL-10 (an anti-inflammatory cytokine) by dendritic cells.
Each microorganism in any environment—including the skin—is in a constant, life-
and-death struggle with other organisms in this environment as they compete for the
available nutrients needed for survival and growth. Microorganisms can make free fatty
acids, enzymes, and virulence factors including toxins, bacteriocins, AMPs, and antibiotics
that help them compete and survive, as illustrated by the following examples. C. acnes,
S. epidermidis, S. aureus, and Malassezia spp. secrete lipases that hydrolyze triglycerides in
sebum to produce free fatty acids that lower the pH of the skin surface and make it more
difficult for non-acid-tolerant microorganisms to grow, and stimulate epithelial cells and
neutrophils to express HBD2, which has antimicrobial activity against Gram-negative
bacteria and Candida spp.84-86 Many fatty acids upregulate AMPs and cytokines in sebaceous
glands, and sebaceous-gland-rich sites have higher levels of AMPs (e.g., psoriasin S100A7,
HBD2, and lipocalin than sebum-poor sites.87-90 Most Corynebacterium spp. contain mycolic
acid in the cell envelope. Mycolic acid can promote γδTcell accumulation with release of
IL-23, an inflammatory cytokine. Some strains of C. acnes produce cutimycin, a thiopeptide
antibiotic that limits S. aureus colonization.89,91 Staphylococcus lugdunensis produces lugdunin,
an antimicrobial peptide that induces keratinocytes to produce AMPs (cathelicidin LL-37
and CXCL8). CXCL8 is a neutrophil chemoattractant produced through the TLR-myeloid
differentiation response protein 88 (TLR-Myd88) pathway.92 S. epidermidis produces
bacteriocins, several types of AMPs, and stimulates keratinocytes and Langerhans cells
to produce AMPs to prevent the growth of other microorganisms. S. epidermidis produces
succinic acid (from fermentation of glycerol) which inhibits some strains of C. acnes.87,93-95
S. epidermidis also produces a serine protease, Esp, that is able to inhibit S. aureus biofilm
formation and colonization.96 Biofilm is involved in the pathogenicity and antimicrobial
susceptibility of C. acnes,97and it is likely that Esp produced by S. epidermidis, or proteases
produced by other microorganisms, may help control biofilm formation of C. acnes, which
in turn decreases C. acnes colonization to decrease the severity of acne.
Skin colonization with commensal microorganisms shapes immunity through activation
of cellular pattern recognition receptors, with distinct activation signatures defining skin
physiology.98 Commensal S. epidermidis, induces IL-17A+ CD8+ T cells that home to the
epidermis, enhance innate barrier immunity, and help prevent pathogen attachment and
colonization.99 Commensals also trigger activation of cellular Toll-like receptors (TLRs)
to enhance immunity and facilitate wound repair.100 Keratinocytes express TLRs and
other immunomodulatory proteins, such as nucleotide-binding oligomerization domain-
containing protein 2 (NOD2), that can recognize pathogenic substances, such as pathogen-
associated molecular patterns including bacterial cell wall peptidoglycans, and stimulate an
immune reaction to them.101,102 Immune response pathways are activated in a TLR-specific
process, which causes the release of cytokines, chemokines, and AMPs to attract circulating
immune cells (e.g., neutrophils and Langerhans cells). Keratinocytes constitutively express
certain AMPs, such as HBD1, while other AMPs are induced in response to injury or
infection.103-105 The Firmicutes and Actinobacteria produce lipoteichoic acid (LTA), which
is a major constituent of Gram-positive bacterial cell walls. LTA is a Toll-like receptor 2
(TLR2) ligand that down-regulates skin inflammation, which in turn, supports barrier
function.82,106
These AMPs help protect the skin from colonization and infection by transient
microorganisms and pathogens.
6. Commensal skin microflora may promote immunological quiescence by up-regulating
production of IL-10 (an anti-inflammatory cytokine) by dendritic cells.
Each microorganism in any environment—including the skin—is in a constant, life-
and-death struggle with other organisms in this environment as they compete for the
available nutrients needed for survival and growth. Microorganisms can make free fatty
acids, enzymes, and virulence factors including toxins, bacteriocins, AMPs, and antibiotics
that help them compete and survive, as illustrated by the following examples. C. acnes,
S. epidermidis, S. aureus, and Malassezia spp. secrete lipases that hydrolyze triglycerides in
sebum to produce free fatty acids that lower the pH of the skin surface and make it more
difficult for non-acid-tolerant microorganisms to grow, and stimulate epithelial cells and
neutrophils to express HBD2, which has antimicrobial activity against Gram-negative
bacteria and Candida spp.84-86 Many fatty acids upregulate AMPs and cytokines in sebaceous
glands, and sebaceous-gland-rich sites have higher levels of AMPs (e.g., psoriasin S100A7,
HBD2, and lipocalin than sebum-poor sites.87-90 Most Corynebacterium spp. contain mycolic
acid in the cell envelope. Mycolic acid can promote γδTcell accumulation with release of
IL-23, an inflammatory cytokine. Some strains of C. acnes produce cutimycin, a thiopeptide
antibiotic that limits S. aureus colonization.89,91 Staphylococcus lugdunensis produces lugdunin,
an antimicrobial peptide that induces keratinocytes to produce AMPs (cathelicidin LL-37
and CXCL8). CXCL8 is a neutrophil chemoattractant produced through the TLR-myeloid
differentiation response protein 88 (TLR-Myd88) pathway.92 S. epidermidis produces
bacteriocins, several types of AMPs, and stimulates keratinocytes and Langerhans cells
to produce AMPs to prevent the growth of other microorganisms. S. epidermidis produces
succinic acid (from fermentation of glycerol) which inhibits some strains of C. acnes.87,93-95
S. epidermidis also produces a serine protease, Esp, that is able to inhibit S. aureus biofilm
formation and colonization.96 Biofilm is involved in the pathogenicity and antimicrobial
susceptibility of C. acnes,97and it is likely that Esp produced by S. epidermidis, or proteases
produced by other microorganisms, may help control biofilm formation of C. acnes, which
in turn decreases C. acnes colonization to decrease the severity of acne.
Skin colonization with commensal microorganisms shapes immunity through activation
of cellular pattern recognition receptors, with distinct activation signatures defining skin
physiology.98 Commensal S. epidermidis, induces IL-17A+ CD8+ T cells that home to the
epidermis, enhance innate barrier immunity, and help prevent pathogen attachment and
colonization.99 Commensals also trigger activation of cellular Toll-like receptors (TLRs)
to enhance immunity and facilitate wound repair.100 Keratinocytes express TLRs and
other immunomodulatory proteins, such as nucleotide-binding oligomerization domain-
containing protein 2 (NOD2), that can recognize pathogenic substances, such as pathogen-
associated molecular patterns including bacterial cell wall peptidoglycans, and stimulate an
immune reaction to them.101,102 Immune response pathways are activated in a TLR-specific
process, which causes the release of cytokines, chemokines, and AMPs to attract circulating
immune cells (e.g., neutrophils and Langerhans cells). Keratinocytes constitutively express
certain AMPs, such as HBD1, while other AMPs are induced in response to injury or
infection.103-105 The Firmicutes and Actinobacteria produce lipoteichoic acid (LTA), which
is a major constituent of Gram-positive bacterial cell walls. LTA is a Toll-like receptor 2
(TLR2) ligand that down-regulates skin inflammation, which in turn, supports barrier
function.82,106