380 JOURNAL OF COSMETIC SCIENCE
into the body. The brick and mortar structure of the SC, proposed almost half a century
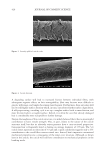
ago,1,14 has been investigated in detail over the years. See Figure 1 for a schematic of the brick
and mortar structure.5 The SC consists of corneocyte bricks embedded in a lipid matrix.3
Covalently bonded lipids attached to the corneocyte envelope compatibilize the brick with
the surrounding lipids. Corneocytes are further attached together by desmosomal proteins.
The proteins within the corneocyte bricks exist as crosslinked keratin and low molecular
weight natural moisturizing factors (NMFs) that help hold water within the SC.
The SC, which was thought to be a dead layer of skin about half a century ago, is now
recognized to be a hot bed of enzyme activity and biological processes.3,5,15 One such
process is the desquamation of skin layers in a layer-by-layer fashion, losing at least one
layer every day, with a fresh layer being exposed regularly. The overall corneum turn-over
period in normal skin is about four weeks. For this to happen, the desmosomal linkages
need to be degraded as the SC layers move to the surface, and this is accomplished by
proteolytic enzymes in the SC. The other two important enzyme driven processes in the
epidermis include the breakdown of filaggrin protein into NMFs and the conversion of
glucosylceramides into ceramides, a key lipid involved in the matrix bilayer lipids. Both
skin pH and hydration affect these enzymatic processes.
The matrix lipids that surround the corneocytes consist of ceramides, cholesterol, and
fatty acids—approximately in a 1:1:1 molar ratio.4 The composition and the possible
structural organization of these bilayer lipids have been discussed in detail over the past
three decades.4,16–18 Ceramides are two-tailed lipids, and depending upon the nature of the
headgroup and the tails, 20 classes of ceramides have been identified in the human SC.15,19
Similarly, fatty acids have also been found with chain lengths, mainly from C20 to C28,
and even low levels of C32.20 Bouwstra and coworkers have investigated the organization
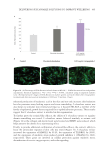
Figure 1. Refined “bricks and mortar” representation of the structural components of the SC. Reproduced
with permission from A. V. Rawlings and R. Voegeli, Stratum corneum proteases and dry skin conditions,
Cell Tissue Res (2013) 351:217–235, DOI 10.1007/s00441-012-1501-x.5
into the body. The brick and mortar structure of the SC, proposed almost half a century
ago,1,14 has been investigated in detail over the years. See Figure 1 for a schematic of the brick
and mortar structure.5 The SC consists of corneocyte bricks embedded in a lipid matrix.3
Covalently bonded lipids attached to the corneocyte envelope compatibilize the brick with
the surrounding lipids. Corneocytes are further attached together by desmosomal proteins.
The proteins within the corneocyte bricks exist as crosslinked keratin and low molecular
weight natural moisturizing factors (NMFs) that help hold water within the SC.
The SC, which was thought to be a dead layer of skin about half a century ago, is now
recognized to be a hot bed of enzyme activity and biological processes.3,5,15 One such
process is the desquamation of skin layers in a layer-by-layer fashion, losing at least one
layer every day, with a fresh layer being exposed regularly. The overall corneum turn-over
period in normal skin is about four weeks. For this to happen, the desmosomal linkages
need to be degraded as the SC layers move to the surface, and this is accomplished by
proteolytic enzymes in the SC. The other two important enzyme driven processes in the
epidermis include the breakdown of filaggrin protein into NMFs and the conversion of
glucosylceramides into ceramides, a key lipid involved in the matrix bilayer lipids. Both
skin pH and hydration affect these enzymatic processes.
The matrix lipids that surround the corneocytes consist of ceramides, cholesterol, and
fatty acids—approximately in a 1:1:1 molar ratio.4 The composition and the possible
structural organization of these bilayer lipids have been discussed in detail over the past
three decades.4,16–18 Ceramides are two-tailed lipids, and depending upon the nature of the
headgroup and the tails, 20 classes of ceramides have been identified in the human SC.15,19
Similarly, fatty acids have also been found with chain lengths, mainly from C20 to C28,
and even low levels of C32.20 Bouwstra and coworkers have investigated the organization
Figure 1. Refined “bricks and mortar” representation of the structural components of the SC. Reproduced
with permission from A. V. Rawlings and R. Voegeli, Stratum corneum proteases and dry skin conditions,
Cell Tissue Res (2013) 351:217–235, DOI 10.1007/s00441-012-1501-x.5