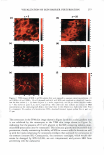
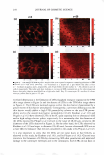
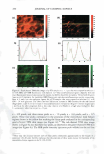
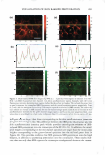
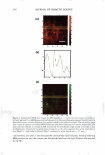
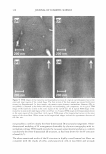
HAIR CURVATURE IN JAPANESE WOMEN 329 suggest the existence of at least two types of cortical cells in curved human hair. Swift (2) reported that straight Mongolian hair shows mainly para-type cortical cells, while curly Caucasian hair has a small amount of the ortho-type cells and very curly Nigerian hair is asymmetrically divided, roughly between 50% each of the ortho- and para-type cortical cells. Kajiura et al. (3) reported that the structural inhomogeneity of IF arrange ment is universally observed in curved human hair (African-American, Caucasian, and Japanese) and they concluded that the structural inhomogeneity may depend on the asymmetrical distribution of the ortho- and para-type cortical cells. These results induce one question: whether hair curvature correlates with the amount of the ortho-type cells or with the asymmetricity of the distribution of the ortho- and para-type cells. Regard ing this question, there is an interesting theory (13) to explain the reason for curved hair shape. When the cortical cell is dehydrated during keratinization, lateral contraction of mac rofibrils in the cortical cell is possibly caused by the removal of water molecules. In such a case, the longitudinal length of the macrofibrils also can be affected. Munro and Carnaby ( 13) proposed a theoretical model to consider the effect of dehydration on the longitudinal length of macrofibrils. They assumed that the IFs are rigid and elastic but free to slide relative to one another. When the IF is aligned parallel to the fiber axis (as in paracortical cells), the longitudinal length of the macrofibril cannot change as a result of the dehydration because of the rigid structure of the standing IF. On the other hand, when the IF is arranged helically (as in orthocortical cells), the longitudinal extension of the macrofibril is possibly caused by the dehydration because the helical angle (direction) of the IF is possibly changed to the direction of fiber axis by the lateral contraction caused by the dehydration. If these two types of macrofibrils (or cortical cells) were laterally attached to each other, to form a complex, the shape of the complex would be deformed by the asymmetric change in the longitudinal lengths of the two types of macrofibrils caused by the dehydration, and the deformation depends on the contents and distribution of those two types of macrofibrils. Munro and Carnaby concluded that for a fiber to have curvature, both of the cortical cell types must be asymmetrically present in the cross section, and the curvature is maximized when there is half of each. These theoretical considerations suggest that hair curvature should correlate with the amount of the ortho-type cells and with the asymmetricity of the distribution of both types of cell. AMINO ACID COMPOSITION OF INNER- AND OUTER-FIBER HALVES The TEM observations of typical highly curved Japanese hair revealed structural differ ences between the outer and inner-fiber halves and thus a structural similarity of curved human hair to crimped wool fibers. These results suggest differences between the protein composition in the outer- and the inner-fiber regions of curls. In the case of ortho- and paracortical cells in wool fiber, the differences in the amino acid composition have been reported (14-16). Hence, we attempted to analyze the differences in amino acid com position between the outer and the inner halves of the curved shape. Typical highly curved hair fibers were divided into the inner and outer halves of the curved shape, and the amino acid composition of each half was determined. The results of the amino acid analyses for the curved human hair are summarized in Table I. The average data of the ortho- and paracortical cells in wool fiber, quoted from three

330 JOURNAL OF COSMETIC SCIENCE Table I Amino Acid Compositions for the Outer and Inner Halves of Curved Japanese Hair Fibers and for the Ortho- and Paracortical Cells of Wool Fibers (moles percent) Curved human hair Wool (Present work) (Average data of refs. 14-16) Amino acids Outer Inner �t P+ Ortho Para �t Asp 5.7 5.5 0.2 ** 6.7 6.2 0.5 Thr 6.9 6.8 0.1 *** 6.4 7.2 -0.8 Ser 11.8 11.8 0.0 10.3 10.6 -0.3 Glu 13.7 13.3 0.4 ** 13.1 12.7 0.4 Pro 7.6 7.8 -0.2 6.8 7.7 -0.9 Gly 5.7 5.6 0.1 * 8.2 7.4 0.8 Ala 4.6 4.6 0.0 5.8 5.6 0.2 Cys 15.8 17.0 -1.2 *** 9.8 11.4 -1.6 Val 5.6 5.5 0.1 5.7 6.0 -0.3 Met 0.4 0.4 0.0 0.4 0.3 0.1 Ile 2.8 2.8 0.0 3.4 3.5 -0.1 Leu 6.4 6.3 0.1 * 7.8 7.1 0.7 Tyr 1.9 1.8 0.1 2.8 2.5 0.3 Phe 1.7 1.6 0.1 2.8 2.5 0.3 Lys 2.6 2.6 0.0 2.7 2.4 0.3 His 0.9 0.9 0.0 0.6 0.5 0.1 Arg 5.9 5.9 0.0 6.9 6.4 0.5 t � = (outer - inner) or (ortho - para). + p means p-values obtained from the paired Student's t-test between "outer" and "inner" *p 0.05 **p 0.01 ***p 0.001. references (14-16), are also shown in the same table for comparison. In these references, the ortho- and paracortical cells of wool fiber were separated and applied on amino acid analyses. Differences in each amino acid between the outer and inner halves and between the ortho- and paracortical cells are shown as "Ll" in this table. The p-values obtained by the paired Student's t-test are summarized as "p" for the curved human hair. The differences (Ll) in amino acids between the outer and inner halves of curved human hair were relatively small, compared with the differences between ortho- and paracortical cells of wool fiber, as shown in Table I. In the case of wool fiber, the analyzed ortho- and paracortical cell samples were separated and purified before amino acid analyses. On the other hand, the divided half fiber of curved human hair is considered to be a mixture of at least two types of cortical cells and cuticle cells, but the major components of the outer- and inner-half fibers are probably ortho- and para-like cortical cells, respectively. The purity of each cell type in each half fiber was, therefore, possibly lower than that of each cell sample of wool. The lower purity of the cell type in the divided human hair is possibly the reason for the smaller differences in the amino acids between the outer and inner halves of curved human hair. Although the differences in curved human hair are smaller than those in wool, we compared the tendency of the differences in curved human hair and wool fibers. In Table I, the outer-fiber half of curved human hair shows the tendencies of more aspartic acid, threonine, glutamic acid, glycine, valine, leucine, tyrosine, and phenylal anine less praline and much less cysteine. Interestingly, these tendencies of curved
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)