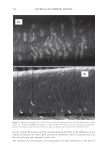
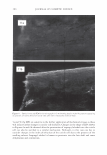
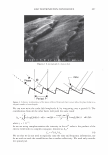
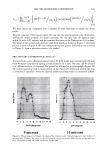
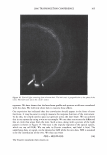
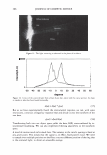
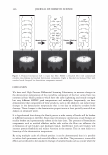
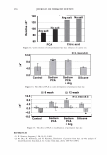
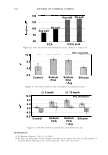
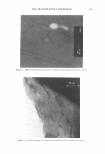
366 JOURNAL OF COSMETIC SCIENCE 9000 8000 a. $ � - 7000 ::::J 6000 5000 1 4000 3000 Figure 10. Modulus data for unmedullated (white box) and medullated hair (gray box). Box plots were obtained from 40 hair fiber measurements from each sample. terizes the yield region. Once the post-yield region is reached, the remaining 70% of the a-helices unfold irreversibly by a compulsory breakdown of the disulphide bonds and they constitute the Y zone. The series zone concept was modified (26) with an extended two phase model to explain the increase of stress in the post-yield region. This model suggests that the increased stress is produced by globular matrix proteins jamming the microfibrils as the fiber is extended into the post-yield region. In this model, water molecules are supposed to be ejected from the matrix at high stress levels, which leads to matrix protein compression between the microfibrils (14). In Chapman's model (27) matrix proteins are supposed to be covalently bonded to fundamental repeat units aligned along the microfibril. The stress-strain curve of the fiber reflects the permanent interaction between microfibril and matrix. Considering TEM observations for medulla, there are several reasons for expecting different mechanical behavior for medullated hairs. First, the microfibrils are randomly distributed through a 20% of the diameter of the fiber. Thus, most of them are not aligned in the axial direction and this could affect the yield region. According to Clement et al. (22), the structures in medulla are cross-linked by peptide bonds from citrulline residues. They might not have the same tensile resistance as the disulfide bonds of the matrix. But when the data is displayed in box plots (Figures 7-10) it is noted that average values are the similar for medullated and unmedullated hair in most cases but that medullated hair shows broader distributions. Considering the data of Figure 7, unmedullated hair has about 5 7 µm of diameter or "pure cortex". On the other hand, medullated hair has about 78 µm of diameter but has a porous inner structure of about 15 µm. Deducting the diameter of medulla, medullated hair has about 63 µm of diameter of "pure cortex". This means that both types of hair should show similar mechanical behaviors. The higher variance in stress-strain curves of medullated hair could be caused by the difference between the fiber diameter minus the medulla diameter. Also, the cavities in the medulla structure as mechanical defects generating cracks which lead to ultimate failure. CONCLUSIONS TEM micrographs allowed the observation of cortical cells disposed randomly in the medulla. The main evidence was the observation of microfibrils.

2006 TRI/PRINCETON CONFERENCE 367 Porous medulla leaves hair less uniform leading to broader distributions of tensile mechanical properties. The cavities of the medulla could act as defects which affect the mechanical properties but do not interfere in the crystalline character of the fiber. ACKNOWLEDGMENTS The authors acknowledge financial support of FAPESP (Funda�ao de Amparo a Pesquisa do Estado de Sao Paulo, Grant 04/13066-0) and of CNPq (Conselho Nacional de Pesquisa, Grant 471061/2004-2). RDCW acknowledges a FAPESP fellowship (Grant 03/13436-0). REFERENCES (1) J. R. Smith, A quantitative method for analyzing AFM images of the outer surfaces of human hair,]. Microsc., 191, 223-228 (1998). (2) J. A. Swift and J. R. Smith, Atomic force microscopy of human hair, Scanning, 22, 310-318 (2000). (3) P. Hallegot, R. Peteranderl, and C. Lechene, In-situ imaging mass spectrometry analysis of melanin granules in the human hair shaft,]. Invest. Dermatol, 122, 381-386 (2004). (4) J. R. Smith and J. A. Swift, Maple syrup urine disease hair reveals the importance of 18- methyleicosanoic acid in cuticular delamination, Micron, 36(3), 261-266 (2005). (5) I. M. Kempson, W. M. Skinner, and P. K. Kirkbride, Calcium distributions in human hair by TOF- SIMS, Biochim. Biophys. Acta, 1624, 1-5 (2003). (6) L. N. Jones, M. Simon, N. R. Watts, F. P. Booy, A. C. Steven and D. A. D. Parry, Intermediate filament structure: hard a-keratin, Biophys. Chem., 68, 83-93 (1997). (7) S. Naito, K. Arai, M. Hirano, N. Nagasawa, and M. J. Sakamoto, Crosslinking structure of keratin. V. Number and type of crosslinks in microstructures of untreated and potassium cyanide treated human hair, Appl. Polym. Sci., 61, 1913-1925 0996). (8) S. Naito and K. Arai, Type and location of SS linkages in human hair and their relation to fiber properties in water,]. Appl. Polym. Sci., 61, 1918-2113 (1996). (9) J. R. Smith and J. A. Swift, Lamellar subcomponents of the cuticular cell membrane complex of mammalian keratin fibres show friction and hardness contrast by AFM, J. Microsc., 206, 182-193 (2002). (10) K. Roper, J. Fohles, D. Peters, and H. Zahn, Morfological composition of the cuticle from chemically treated wool. 1. Calculating endocuticle content in isolated cuticle from results of amino-acid analysis, Text. Res.]., 54, 139-143 (1984). (11) E. G. Bendit and M. Feughelman (Eds.), "Keratin," Encyclopedia of Polymer Science and Technology, 8th Ed., Chapter 1 (1967). (12) A. Kelch, S. Wessel, T. Will, R. Hintze, R. Wepf, and R. Wiesendanger, Penetration pathways of fluorescent dyes in human hair fibres investigated by scanning near-field optical microscopy, J. Microsc., 200(3), 179-186 (2000). (13) L. Pi.ietsh and M. R. Moeller, On pathways for small molecules into and out of human hair fibers,]. Forensic Sci., 41(1), 121-125 (1996). (14) L. Kreplak, A. Franbourg, F. Briki, F. Leroy, D. Dalle J., and Doucet, A new deformation model of hard alpha-keratin fibers at the nanometer sacale: implications for hard alpha-keratin intermediate filament mechanical properties, Biophys. ]., 82, 2265-2274 (2002). (15) D. W. Deedrick, Microscopy of Hair Part 1: A Practical Guide and Manual for Human Hairs, Forensic Sci. Commun., 1(6) (2004). (16) L. Kreplak, F. Briki, Y. Duvault, J. Doucet, C. Merigoux, F. Leroy, L. Leveque, L. Miller, G. L. Carr, G. P. Williams, and P. Dumas, Profiling lipids across Caucasian and Afro-American hair transverse cuts, using synchroton infrared microspectrometry, Int. J. Cosmet. Sci., 23, 369-374 (2001). (17) G. Mahrle and C. E. Orfanos, The spongious keratin and the medulla of human scalp hair, Arch. Derm. Res., 241, 305-316 (1971). (18) S. Nagase, S. Shibuichi, K. Ando, E. Kari ya, and N. Satoh, Influence of internal structures of hair fiber
Purchased for the exclusive use of nofirst nolast (unknown) From: SCC Media Library & Resource Center (library.scconline.org)